
Нагревательные индукционные установки | ПЕТРА
Индуктор изолирован от сети и заземлён
Широкий набор индукторов без переключений внутри установки
Эффективная работа на пайке и поверхностной закалке без переключений внутри установки
Нагревательный пост легко встраивается в технологическую линию
Ссылки
Индукционная пайка резцов, фрез и другого металлорежущего инструмента
ТВЧ закалка валов
ТВЧ закалка крупномодульных шестерен по впадине
ТВЧ закалка шестерен в кольцевом индукторе
Нагрев кузнечных заготовок для штамповочного производства
Индукционная пайка пакетов роторов центробежных погружных насосов
Область применения
Установки индукционного нагрева ТВЧ ПЕТРА-0501 используются во многих отраслях промышленности для осуществления эффективных технологических процессов электротермической обработки изделий. Установки ТВЧ применяются там, где требуется быстрый бесконтактный нагрев металлов и других проводящих материалов. В зависимости от исполнения по номинальной частоте и мощности индукционные установки ТВЧ ПЕТРА-0501 применяются в следующих областях:
В зависимости от исполнения по номинальной частоте и мощности индукционные установки ТВЧ ПЕТРА-0501 применяются в следующих областях:
Описание
Индукционный нагреватель металла ПЕТРА-0393 оснащён высокочастотным закалочным трансформатором с сердечником из феррита или аморфного железа. Органы переключения коэффициента трансформации вынесены на внешние панели нагревательного поста. Переключатель первичной обмотки имеет два положения: «ПАЙКА» и «ЗАКАЛКА». Режим «ПАЙКА» применяется, когда необходим плавный, сквозной индукционный нагрев заготовки. Допускается работа на заполненный, полупустой и пустой индуктор. Режим «ЗАКАЛКА» применяется, если необходимо полную мощность установки выделить в поверхностном слое заготовки, не нагрев при этом сердцевину. Предусмотрена возможность изменять число используемых витков вторичной обмотки трансформатора от 1 до 3. Благодаря этому, без каких-либо переключений внутри установки к ней могут подключаться индукторы с индуктивностью от 0,2 до 1,8 мкГн. Важно, что при этом индукционная установка способна обеспечивать номинальную или близкую к номинальной мощность в индукторе.
Органы переключения коэффициента трансформации вынесены на внешние панели нагревательного поста. Переключатель первичной обмотки имеет два положения: «ПАЙКА» и «ЗАКАЛКА». Режим «ПАЙКА» применяется, когда необходим плавный, сквозной индукционный нагрев заготовки. Допускается работа на заполненный, полупустой и пустой индуктор. Режим «ЗАКАЛКА» применяется, если необходимо полную мощность установки выделить в поверхностном слое заготовки, не нагрев при этом сердцевину. Предусмотрена возможность изменять число используемых витков вторичной обмотки трансформатора от 1 до 3. Благодаря этому, без каких-либо переключений внутри установки к ней могут подключаться индукторы с индуктивностью от 0,2 до 1,8 мкГн. Важно, что при этом индукционная установка способна обеспечивать номинальную или близкую к номинальной мощность в индукторе.
Питание индукционного нагревательного поста осуществляется от преобразователя частоты ПЕТРА-0132 или ПЕТРА-0133. Система автоматического управления преобразователей позволяет стабилизировать напряжения индуктора или мощность, потребляемую из сети, на заданном уровне. Преобразователи имеют разветвлённую систему защит и блокировок.
Преобразователи имеют разветвлённую систему защит и блокировок.
Охлаждение установки производится технической цеховой водой. Индукционная установки ТВЧ ПЕТРА-0501 с преобразователями ПЕТРА-0132 комплектуются теплообменными станциями ПЕТРА-0395 для двухконтурного охлаждения. В случае комплектации установки преобразователем ПЕТРА-0133 (60-100 кВт, 22.0 кГц) теплообменная станция встроена в преобразователь. Поэтому отдельной теплообменной станции не требуется. По двухконтурной схеме (дистиллированной водой) охлаждается преобразователь частоты и нагревательный блок. Индуктор охлаждается напрямую технической водой. Для подключения охлаждения индуктора на блоке нагревательном предусмотрены отдельные вводы для технической воды.
Установка индукционная нагревательная ПЕТРА-0501 эффективно заменяет собой ламповые ТВЧ-установки типа ЛЗ и ВЧГ. По сравнению с ламповыми установками ПЕТРА-0501 имеет высокий КПД, малые габариты и не требует дополнительного времени перед началом и по окончании работы на разогрев и остывание лампы.
Отличительная особенность индукционной установки ПЕТРА-0501 – высокая допустимая полная мощность индуктора и возможность обеспечения нагрева в индукторах с малым cosφ.
Технические характеристики
* Параметры выпускаемого оборудования могут отличаться от табличных. Запрашивайте уточнение при обращении в ООО НКВП «Петра».
| Рпот, кВт | Полная мощность в индукторе, кВА | Fинд, кГц | Сквозной нагрев, Т>750oC | Поверхностная закалка | |||
| Ømin, мм | Ønorm, мм | Ømin, мм | Ømax, мм | Глубина, мм | |||
| 60 | 400, 500 | 44.0 | 6.0 | 20…25 | 8 | 120 | 1.0 … 2.2 |
| 60, 100, 160 | 400, 500, 800 | 22.0 | 8.0 | 20…30 | 11 | 120, 200, 250 | 1. 6…4.0 6…4.0 |
| 100, 160, 250, 320 | 500, 800, 3200 | 8.0 | 15…20 | 45…60 | 18…20 | 200, 250, 300, 500 | 2.75 … 5.5 |
| 100, 160, 250, 320 | 800, 3200 | 2.4 | 25.0 | 60…80 | 35 | 200, 250, 300, 500 | 5.0 … 10.0 |
Габаритный чертеж
Скачать
IGBT-Преобразователи частоты ПЕТРА для индукционного нагрева
Скачать (PDF)
Индукционная нагревательная установка ПЕТРА-0501
Скачать (PDF)
Индукционные плавильные Печи ИСТ
Скачать (PDF)
Функциональное назначение экранов панели индикации преобразователей частоты ПЕТРА
Скачать (PDF)
Пусконаладочные работы
Подготовка пусконаладочных работ начинается на этапе заключения договора. Специалисты ООО НКВП «Петра» согласовывают план размещения оборудования в производственных условиях покупателя, подвод коммуникаций (электроэнергия, вода, канализация и др. ), сопряжение с оборудованием покупателя.
), сопряжение с оборудованием покупателя.
Пусконаладочные работы включают в себя: выезд специалиста к Заказчику; подключение и настройку оборудования для обеспечения технологического режима; обучение производственного и обслуживающего персонала. Стоимость работ составляет 5…10% от цены поставленного оборудования.
Срок изготовления
Срок изготовления зависит от готовности оборудования на сборочном производстве и сложности Вашего заказа. Поэтому уточняйте срок изготовления при каждом обращении к нашим менеджерам. Обычно срок поставки не превышает: для преобразователей частоты – 2-х месяцев, для индукционных установок – 3-х месяцев.
Доставка
Доставка оборудования производится самовывозом на транспорте покупателя или транспортной компанией.
Условия оплаты
Как правило, применяется поэтапная оплата по формуле: 50% — аванс, 50% — оплата перед отгрузкой оборудования. Другие формы оплаты – по согласованию с покупателем.
Гарантия
В случае возникновения гарантийного случая в период гарантийного обслуживания работы по ремонту оборудования производятся бесплатно. Срок действия гарантии — 12 месяцев со дня ввода в эксплуатацию. Возможны другие условия гарантии, которые предусматриваются договором поставки.
Срок действия гарантии — 12 месяцев со дня ввода в эксплуатацию. Возможны другие условия гарантии, которые предусматриваются договором поставки.
Индукционные установки – Индукционная литейная установка – Установка индукционного нагрева
ИНДУКЦИOННAЯ УСТАНOВКA — это оборудование, необходимое для индукционного нагрева металлических изделий и деталей электрическим током, индуцируемым переменным магнитным полем. РОСИНДУКТОР — это установка индукционного нагрева от профессионалов. Здесь можно купить по хорошей цене установки для закалки, пайки и плавки. Каждая индукционная литейная установка собрана по схемам с использованием современных IGBT модулей. Специалисты нашей компании модернизируют индукционные установки, а так же их ремонт и обслуживание.
- Установка индукционного нагрева
- Индукционная литейная установка
- Преимущества индукционных установок
- Принцип работы индукционных установок
- Применение индукционных установок
Установки индукционного нагрева делятся на транзисторные и тиристорные. Тиристорная индукционная установка нагрева имеет слишком узкий предел изменения генерируемой частоты. Транзисторные установки индукционного нагрева металла отличаются более высоким КПД, они не нуждаются в предварительном прогреве. Кроме того каждая установка нагрева имеет контроль и регулировку выходной частоты в большом диапазоне.
Тиристорная индукционная установка нагрева имеет слишком узкий предел изменения генерируемой частоты. Транзисторные установки индукционного нагрева металла отличаются более высоким КПД, они не нуждаются в предварительном прогреве. Кроме того каждая установка нагрева имеет контроль и регулировку выходной частоты в большом диапазоне.
- Новосибирск
- Новосибирск
Высокочастотный индукционный нагреватель 100 кВт
- Москва
Среднечастотный индукционный нагреватель мощностью 15 кВт
- Новосибирск
Высокочастотный индукционный нагреватель мощностью 100 кВт
- Пермь
Высокочастотный индукционный нагреватель мощностью 60 кВт
- Екатеринбург
Среднечастотный индукционный нагреватель мощностью 80 кВт
- Красноярск
Высокочастотный индукционный нагреватель мощностью 60 кВт
- Челябинск
Среднечастотный индукционный нагреватель мощностью 60 кВт
- Самара
Высокочастотный индукционный нагреватель мощностью 160 кВт
- Калуга
Высокочастотный индукционный нагреватель мощностью 60 кВт
- Екатеринбург Высокочастотный индукционный нагреватель мощностью 120 кВт
- Екатеринбург
Среднечастотный индукционный нагреватель мощностью 120 кВт
Установки индукционного нагрева (УИН) делятся на транзисторные и тиристорные.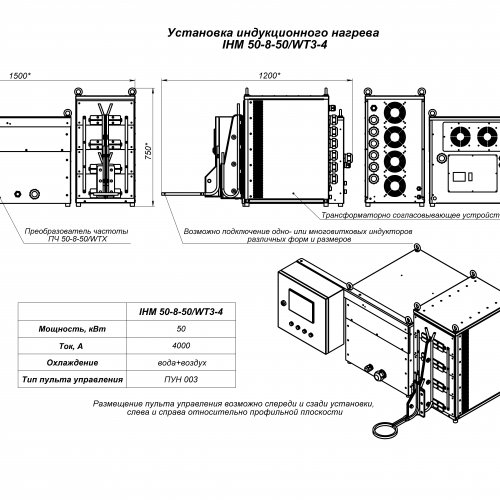 Тиристорная индукционная нагревательная установка имеет слишком узкий предел изменения генерируемой частоты. Транзисторные установки индукционного нагрева металла отличаются более высоким КПД, они не нуждаются в предварительном прогреве. Кроме того транзисторные ТВЧ генераторы подходят для контроля и регулировки выходной частоты в большом диапазоне.
Тиристорная индукционная нагревательная установка имеет слишком узкий предел изменения генерируемой частоты. Транзисторные установки индукционного нагрева металла отличаются более высоким КПД, они не нуждаются в предварительном прогреве. Кроме того транзисторные ТВЧ генераторы подходят для контроля и регулировки выходной частоты в большом диапазоне.
Установки индукционного нагрева бывают следующих типов:
- Среднечастотные индукционные установки, работающие в частотном диапазоне от 0,5 до 20 кГц и имеющие мощность от 15 до 500 кВт, идеально подходят для максимально глубокого прогрева металлических деталей. Глубина прогрева может достигать 10 мм. Этот вид установок применяется для горячей штамповки, плавки черных и цветных металлов и глубокой закалки металлов.
- Высокочастотные индукционные установки осуществляют работу с частотой от 30 до 100 кГц. Глубина прогрева деталей в этих установках составляет всего 2-3 мм, благодаря чему они подходят для поверхностной закалки металлических изделий, восстановления деталей, сварки и наплавки деталей.

- Кузнечные индукционные установки необходимы для эффективного нагрева металлических заготовок. Используемая температура колеблется от 500 до 1200 градусов в зависимости от вида металла и следующих за нагревом процессов. Так, перед горячей штамповкой алюминиевые заготовки необходимо нагреть до 500 градусов, а медные — до 700.
- Индукционные плавильные установки необходимы для плавки цветных и черных металлов. Такие установки оборудуются индукционными печами, подходящими для обработки того или иного металла или сплава.
Индукционная литейная установка
Индукционные литейные установки рассчитаны для работы со сплавом неблагородных и благородных металлов. В таких установках используются печи с тиглями, наиболее подходящими для определенной шихты. Индукционные литейные печи обладают высокой производительностью и легко встраиваются в рабочий конвейер цеха.
Применение индукционных установок
Индукционные установки широко применяются в чугунном и сталелитейном производстве, а также в ювелирном и любом другом производстве, связанным с обработкой металла. Разные индукционные установки используются для поверхностной и глубокой закалки металлических деталей, наплавки, пайки, плавки, сварки металлов, гибки и предварительного нагрева металла, стерилизации медицинских инструментов, ювелирного производства.
Разные индукционные установки используются для поверхностной и глубокой закалки металлических деталей, наплавки, пайки, плавки, сварки металлов, гибки и предварительного нагрева металла, стерилизации медицинских инструментов, ювелирного производства.
Принцип работы индукционных установок
Принцип работы индукционных установок основан на нагреве металлических материалов электрическими токами, которые создаются переменным магнитным полем индуктора. Индуктор представляет собой несколько витков провода. При помощи генератора в индукторе создаются мощные токи, благодаря чему образуется магнитное поле, которое наводит вихревые токи в помещенной в центр индуктора заготовке. В результате происходит нагрев заготовки под действием закона Джоуля-Ленца. Интенсивность и глубина прогрева металлических деталей зависит от выбранной частоты.
Преимущества индукционных установок
Преимущества индукционных установок очевидны: высокий уровень КПД, высокая скорость прогрева или плавления электропроводящего материала, возможность проведения местного нагрева, отсутствие загрязнения материала продуктами горения, нагрев металлических деталей через стенки камеры, выполненной из материалов, не нагревающихся вследствие электромагнитного излучения, возможность автоматизации большинства процессов, происходящих в установке.
Нужна более подробная информация на индукционные установки?
Москва + 7 (499) 649-29-80
Санкт-Петербург + 7 (812) 426-33-17
Челябинск + 7 (351) 729-83-71
Разгадка размножения растений путем гаплоидной индукции для инноваций в селекции растений
Forster, B.P. & Thomas, WTB в Plant Breeding Reviews (Ed. Janick, J.) 57–88 (John Wiley & Sons, 2005).
Wędzony, M. et al. в Advances in Haploid Production in High Plants (под редакцией Touraev, A. et al.) 1–33 (Springer, 2009).
Жиль, Л. М., Мартинан, Ж.-П., Роговский, П. М. и Видиз, Т. Индукция гаплоидов у растений. Курс. биол. 27 , R1095–R1097 (2017).
Артикул КАС пабмед Google ученый
Производство удвоенных гаплоидов в культуре Растения: Руководство (под редакцией Малушински, М. и др.) (Springer, 2003).
и др.) (Springer, 2003).
Артикул КАС пабмед Google ученый
Форстер Б.П., Хеберле-Борс Э., Каша К.Дж. и Тураев А. Возрождение гаплоидов у высших растений. Trends Plant Sci. 12 , 368–375 (2007).
Артикул КАС пабмед Google ученый
Guha, S. & Maheshwari, S.C. Получение in vitro эмбрионов из пыльников Datura . Природа 204 , 497–497 (1964).
Достижения в производстве гаплоидов Высшее Растения (под редакцией Тураева, А. и др.) (Springer, 2009).
Гаплоиды в Улучшение урожая II (ред. Палмер, К.Э.Д. и др.) (Springer, 2005).
Bajaj, YPS Гаплоиды в улучшении сельскохозяйственных культур I Vol. 12 (Спрингер-Верлаг, 1990).
Продуцирование гаплоидов in vitro в высших растениях (под редакцией Jain, S. et al.) Vol. 25 (Спрингер, 1996).
Исии Т., Карими-Аштияни Р. и Хоубен А. Гаплоидизация путем удаления хромосом: средства и механизмы. год. Преподобный завод биол. 67 , 421–438 (2016).
Артикул КАС пабмед Google ученый
Kalinowska, K. et al. Современные и новые разработки гаплоидных технологий in vivo. Теор. заявл. Жене. 132 , 593–605 (2019).
Артикул КАС пабмед Google ученый
Кермикл, Дж. Л. Андрогенез, обусловленный мутацией кукурузы.
Артикул КАС пабмед Google ученый
Coe, E.H. Линия кукурузы с высокой гаплоидной частотой. Американский натуралист 93 , 381–382 (1959).
Артикул Google ученый
Робер, Ф.К., Гордильо, Г.А. и Гейгер, Х.Х. Индукция гаплоидов in vivo у кукурузы: эффективность новых индукторов и значение удвоенных гаплоидных линий в селекции гибридов. Maydica 50 , 275 (2005).
Google ученый
Гейгер, Х. Х. и Гордильо, Г. А. Удвоенные гаплоиды в селекции гибридов кукурузы. Maydica 54 , 485–499 (2009).
Google ученый
Саркар, К. Р. и Коу, Э. Х. Генетический анализ происхождения материнских гаплоидов у кукурузы.
КАС пабмед ПабМед Центральный Google ученый
Barret, P., Brinkmann, M. & Beckert, M. Главный локус, экспрессированный в мужском гаметофите с неполной пенетрантностью, отвечает за гиногенез in situ у кукурузы. Теор. заявл. Жене. 117 , 581–594 (2008).
Артикул КАС пабмед Google ученый
Hu, H. et al. Генетическая основа индукции гаплоидов у кукурузы идентифицирована с помощью нового метода полногеномной ассоциации. Генетика 202 , 1267–1276 (2016).
Артикул КАС пабмед ПабМед Центральный Google ученый
Пригге, В. и др. Новое понимание генетики индукции материнских гаплоидов in vivo, основы технологии двойных гаплоидов у кукурузы.
Артикул КАС пабмед ПабМед Центральный Google ученый
Gilles, L.M. et al. Потеря специфичной для пыльцы фосфолипазы NOT LIKE DAD запускает гиногенез у кукурузы. EMBO J. 36 , 707–717 (2017).
Артикул КАС пабмед ПабМед Центральный Google ученый
Kelliher, T. et al. MATRILINEAL, специфичная для спермы фосфолипаза, запускает гаплоидную индукцию кукурузы. Природа 542 , 105–109 (2017).
Артикул КАС пабмед Google ученый
Лю, К. и др. Вставка 4 п.н. в ZmPLA1, кодирующая предполагаемую фосфолипазу а, вызывает гаплоидную индукцию у кукурузы. Мол. Завод 10 , 520–522 (2017).
Артикул КАС пабмед Google ученый
Zhong, Y. et al. Мутация ZmDMP усиливает индукцию гаплоидов у кукурузы. Нац. Растения 5 , 575–580 (2019).
Артикул пабмед Google ученый
Ravi, M. & Chan, S.W.L. Гаплоидные растения, полученные методом элиминации генома, опосредованной центромерами. Природа 464 , 615–618 (2010).
Артикул КАС пабмед Google ученый
Tan, E.H. et al. Катастрофическая хромосомная перестройка при элиминации генома у растений. eLife 4 , e06516 (2015).
Артикул пабмед ПабМед Центральный КАС Google ученый
Kelliher, T. et al. Материнские гаплоиды предпочтительно индуцируются трансгенной комплементацией CENh4-tailswap у кукурузы. Фронт. Растениевод. 7 , 414 (2016).
Артикул пабмед ПабМед Центральный Google ученый
Махешвари, С. и др. Естественные различия в CENh4 влияют на сегрегацию хромосом в зиготических митозах гибридов. Генетика PLoS. 11 , e1004970 (2015).
Артикул пабмед ПабМед Центральный КАС Google ученый
Карими-Аштияни, Р. и др. Точечная мутация нарушает центромерную загрузку CENh4 и индуцирует гаплоидные растения. Проц. Натл акад. науч. США 112 , 11211–11216 (2015 г.).
Артикул КАС пабмед Google ученый
Куппу, С. и др. Точковые мутации в центромерном гистоне вызывают постзиготную несовместимость и однородительское наследование. Генетика PLoS. 11 , е1005494 (2015).
Артикул пабмед ПабМед Центральный КАС Google ученый
Бритт, А. Б. и Куппу, С. Cenh4: новый игрок в технологии гаплоидной индукции. Фронт. Растениевод. 7 , 357 (2016).
Артикул пабмед ПабМед Центральный Google ученый
Куппу, С. и др. Различные изменения, в том числе CRISPR/Cas9опосредованные делеции в CENh4 приводят к гаплоидной индукции при ауткроссинге. Завод Биотех. Дж . (В прессе).
van Dun, C., Lelivelt, C. & Movahedi, S. Нетрансгенные гаплоидные индукторные линии в тыквенных культурах. Всемирный патент WO/2017/081009A1 (2017 г.).
Op Den Camp, RHM, Van Dijk, PJ & Gallard, A. Способ получения гаплоидных и последующих удвоенных гаплоидных растений. Всемирный патент WO/2017/200386 (2017 г.).
Бора, А., Джха, У. К., Адхимулам, П., Бишт, Д. и Сингх, Н. П. Цитоплазматическая мужская стерильность (CMS) в гибридной селекции полевых культур. Plant Cell Rep. 35 , 967–993 (2016).
Артикул КАС пабмед Google ученый
Li, L., Xu, X., Jin, W. & Chen, S. Морфологические и молекулярные доказательства интрогрессии ДНК при гаплоидной индукции с помощью индуктора CAUHOI с высоким содержанием масла в кукурузе. Планта 230 , 367–376 (2009).
Артикул КАС пабмед Google ученый
Zhang, Z. et al. Ликвидация хромосом и продукция гаплоидов in vivo , индуцированная индукторной линией Stock 6 , полученной из кукурузы ( Zea mays L.). Plant Cell Rep. 27 , 1851–1860 (2008).
Артикул КАС пабмед Google ученый
Чжао, X., Сюй, X., Се, Х., Чен, С. и Цзинь, В. Оплодотворение и однородительская элиминация хромосом во время скрещиваний с гаплоидными индукторами кукурузы. Завод Физиол. 163 , 721–731 (2013).
Артикул КАС пабмед ПабМед Центральный Google ученый
Цю, Ф. и др. Морфологические, клеточные и молекулярные доказательства случайной элиминации хромосом in vivo при гаплоидной индукции у кукурузы. Курс. биол. растений 1 , 83–90 (2014).
Артикул Google ученый
Wedzony, M., Röber, F. & Geiger, H. Элиминация хромосом, наблюдаемая у самоопыленных потомков индукторной линии кукурузы RWS. в XVII Международный конгресс по репродукции половых растений 173 (2002).
Лю, Л. и др. Гаплоидная индукция in vivo приводит к увеличению частоты двойных эмбрионов и аномальному оплодотворению у кукурузы. BMC Plant Biol . 18 , 313 (2018).
Артикул КАС пабмед ПабМед Центральный Google ученый
Былич В., Чалык С. Существование пыльцевых зерен с парой морфологически различных ядер сперматозоидов как возможная причина гаплоидной индуцирующей способности у линии ЗМС. Информационный бюллетень по сотрудничеству в области генетики кукурузы 70 , 33 (1996).
Google ученый
Чалык С., Бауманн А., Даниэль Г. и Эдер Дж. Анеуплоидия как возможная причина индукции гаплоидов у кукурузы. Информационный бюллетень сотрудничества в области генетики кукурузы 77 , 29–30 (2003).
Google ученый
Тянь, X. и др. Гетерооплодотворение вместе с неудачным слиянием яйцеклеток и сперматозоидов поддерживает одиночное оплодотворение, участвующее в гаплоидной индукции in vivo у кукурузы. Дж. Экспл. Бот . 69 , 4689–4701 (2018).
Артикул КАС пабмед ПабМед Центральный Google ученый
Li, X. et al. Секвенирование одного ядра выявило фрагментацию хромосом сперматид как возможную причину гаплоидной индукции кукурузы. Нац. коммун. 8 , 991 (2017).
Артикул пабмед ПабМед Центральный КАС Google ученый
Swapna, M. & Sarkar, KR. Аномальное оплодотворение в линиях индукторов гаплоидии у кукурузы ( Zea mays L). Maydica 56 , 1717 (2012).
Google ученый
Махендру А. и Саркар К. Р. Цитологический анализ пыльцы линий индукторов гаплоидии кукурузы ( Zea mays L.). Индийский . Ж. Жене. пл. бр. 60 , 37–43 (2000).
Google ученый
Джексон, Д. Без секса, пожалуйста, мы (в) разведении. EMBO J. 36 , 703–704 (2017).
Артикул КАС пабмед ПабМед Центральный Google ученый
Cyprys, P., Lindemeier, M. & Sprunck, S. Слиянию гамет способствуют два мембранных белка DUF679, экспрессируемых сперматозоидами. Нац. Растения 5 , 253–257 (2019).
Артикул КАС пабмед Google ученый
Такахаши Т. и др. Белок мембраны мужской гамет DMP9/DAU2 необходим для двойного оплодотворения у цветковых растений. Разработка 145 , dev170076 (2018).
Лю, К. и др. Точное картирование qhir8 , влияющих на индукцию гаплоидов in vivo у кукурузы. Теор. заявл. Жене. 128 , 2507–2515 (2015).
Артикул КАС пабмед Google ученый
Schlupp, I. Эволюционная экология гиногенеза. год. Преподобный Экол. Эвол. Сист. 36 , 399–417 (2005).
Артикул Google ученый
Уоттс, А., Кумар, В. и Бхат, С. Р. Центромерный гистоновый белок h4: от фундаментальных исследований до применения в селекции растений. J. Plant Biochem. Биотехнолог. 25 , 339–348 (2016).
Артикул КАС Google ученый
Wang, N. & Dawe, R.K. Размер центромер и его связь с образованием гаплоидов у растений. Мол. Завод 11 , 398–406 (2018).
Артикул КАС пабмед Google ученый
Comai, L. & Tan, E.H. Гаплоидная индукция и нестабильность генома. Тенденции Жене. 35 , 791–803 (2019).
Артикул КАС пабмед Google ученый
Саней, М., Пикеринг, Р., Кумке, К., Насуда, С. и Хоубен, А. Потеря центромерного гистона h4 (CENh4) из центромер предшествует однородительской элиминации хромосом у межвидовых гибридов ячменя. Проц. Натл акад. науч. США 108 , E498–E505 (2011 г.).
Артикул КАС пабмед Google ученый
Гао, К. Будущее технологий CRISPR в сельском хозяйстве. Нац. Преподобный Мол. Клеточная биол. 19 , 275–276 (2018).
Артикул КАС пабмед Google ученый
Kelliher, T. et al. Одноэтапное редактирование генома зародышевой плазмы элитных культур во время гаплоидной индукции. Нац. Биотех . 37 , 287–292 (2019).
Артикул КАС Google ученый
Ван, Б. и др. Разработка системы редактирования генома, опосредованной гаплоидным индуктором, для ускорения селекции кукурузы. Мол. Завод 12 , 597–602 (2019).
Артикул пабмед КАС Google ученый
d’Erfurth, I. et al. Превращение мейоза в митоз. PLoS Биол. 7 , e1000124 (2009 г.).
Артикул пабмед ПабМед Центральный КАС Google ученый
Mieulet, D. et al. Превращение мейоза риса в митоз. Сотовые Res. 26 , 1242–1254 (2016).
Артикул КАС пабмед ПабМед Центральный Google ученый
Wang, C. et al. Клональные семена гибридного риса путем одновременной геномной инженерии генов мейоза и оплодотворения. Нац. Биотех . 37 , 283 (2019).
Артикул КАС Google ученый
Qu, Y. et al. Картирование QTL для аборта ядра, вызванного индукцией гаплоидов in vivo у кукурузы ( Zea mays L.). PLoS ONE 15 , e0228411 (2020).
Артикул КАС пабмед ПабМед Центральный Google ученый
Yao, L. et al. Мутация OsMATL индуцирует образование гаплоидных семян у индика-риса. Нац. Растения 4 , 530–533 (2018).
Артикул КАС пабмед Google ученый
Liu, C. et al. Расширение системы гаплоидной индукции in vivo от диплоидной кукурузы до гексаплоидной пшеницы. Завод Биотехнолог. J. 18 , 316–318 (2020).
Артикул пабмед Google ученый
Лю, Х. и др. Эффективная индукция гаплоидных растений у пшеницы путем редактирования TaMTL с использованием оптимизированной системы CRISPR, опосредованной Agrobacterium . Дж. Экспл. Бот . 71 , 1337–1349 (2020).
Артикул пабмед Google ученый
Прасанна, Б., Чайкам, В. и Махуку, Г. Двойная гаплоидная технология в селекции кукурузы: теория и практика (CIMMYT, 2012).
Чайкам В., Гауда М., Наир С.К., Мельчингер А.Е. и Боддупалли П.М. Полногеномное ассоциативное исследование для выявления геномных областей, влияющих на спонтанную фертильность гаплоидов кукурузы. Euphytica 215 , 138 (2019).
Артикул пабмед ПабМед Центральный КАС Google ученый
Ren, J., Wu, P., Tian, X., Lübberstedt, T. & Chen, S. Картирование QTL для гаплоидной мужской фертильности методом искажения сегрегации и точное картирование ключевого QTL qhmf4 в кукуруза. Теор. заявл. Жене. 130 , 1349–1359 (2017).
Артикул КАС пабмед Google ученый
Ян, Дж. и др. Генетический анализ гаплоидной мужской фертильности у кукурузы ( Zea mays L.). Растениеводство 138 , 259–265 (2019).
Артикул КАС Google ученый
Рен, Дж. и др. Картирование QTL и идентификация генов-кандидатов, обеспечивающих спонтанное удвоение гаплоидного генома у кукурузы ( Zea mays L.). Растениевод. 293 , 110337 (2020).
Артикул КАС пабмед Google ученый
Бурман, Н. А., Фрей, Великобритания и Любберштедт, Т. Влияние спонтанного удвоения гаплоидного генома на селекцию кукурузы. Растения 9 , 369 (2020).
Артикул ПабМед Центральный Google ученый
Бегейн Р. Ф., Вангсгаард К., Рулунд Н. и Студер Б. In Разведение в мире Дефицит (ред. Ролдан-Руис, И. и др.) 151–155 (Springer, 2016).
Донг Ю.-К. и другие. Гаплоиды андрогенеза, гиногенеза и партеногенеза у видов тыквенных. Представитель растительных клеток . 35 , 1991–2019 (2016).
Артикул КАС пабмед Google ученый
Hussain, T. & Franks, C. in Sorghum Vol. 1931 г. (редакторы Чжао, З.-Ю. и Дальберг, Дж.) 49–59 (Спрингер, 2019).
Клойбер-Майц М., Викхорст С., Болдуан С. и Узунова М. Гаплоидизация сорго. Европейский патент EP3366778A1 (2018 г.).
Прайминг, индукция и модуляция защитных реакций растений бактериальными липополисахаридами
Обзор
. 2007;13(2):69-84.
дои: 10.1177/0968051
9399.
Мари-Энн Ньюман 1 , Дж. Максвелл Доу, Антонио Молинаро, Микеланджело Паррилли
принадлежность
- 1 Кафедра биологии растений, Факультет наук о жизни, Копенгагенский университет, Фредериксберг, Дания.
 [email protected]
[email protected]
- PMID: 17621548
- DOI:
10.1177/0968051
9399
Обзор
Mari-Anne Newman et al. J Эндотоксин Res. 2007.
. 2007;13(2):69-84.
дои: 10.1177/0968051
9399.
Авторы
Мари-Энн Ньюман 1 , Дж. Максвелл Доу, Антонио Молинаро, Микеланджело Паррилли
принадлежность
- 1 Кафедра биологии растений, Факультет наук о жизни, Копенгагенский университет, Фредериксберг, Дания. [email protected]
- PMID: 17621548
- DOI:
10.1177/0968051
9399
Абстрактный
Бактериальные липополисахариды (ЛПС) играют несколько ролей во взаимодействиях растений и микробов. ЛПС способствует низкой проницаемости внешней мембраны, которая действует как барьер для защиты бактерий от противомикробных веществ растительного происхождения. И наоборот, восприятие LPS растительными клетками может привести к запуску защитных реакций или к праймингу растения, чтобы реагировать более быстро и/или в большей степени на последующее воздействие патогена. ЛПС симбиотических бактерий могут оказывать на растения совершенно иное действие, чем действие патогенов. Появляются некоторые подробности о структурах внутри LPS, которые ответственны за индукцию этих различных реакций растений. Фрагмент липида А не несет единоличной ответственности за все эффекты ЛПС в растениях; основные компоненты олигосахаридов и О-антигена могут вызывать специфические ответы. Здесь мы рассматриваем влияние ЛПС на индукцию связанных с защитой реакций у растений, структуры внутри ЛПС, ответственные за проявление этих эффектов, и обсуждаем возможную природу (пока не идентифицированных) рецепторов ЛПС в растениях.
Похожие статьи
Связанные с микробами молекулярные паттерны врожденного иммунитета: экстракция и химический анализ липополисахаридов грамотрицательных бактерий.
Де Кастро К., Паррилли М., Холст О., Молинаро А. Де Кастро С. и др. Методы Энзимол. 2010; 480:89-115. doi: 10.1016/S0076-6879(10)80005-9. Методы Энзимол. 2010. PMID: 20816206 Обзор.
Роль липополисахаридов в индукции защитных реакций растений.
Эрбс Г., Ньюман М.А. Эрбс Г. и соавт. Мол Плант Патол. 2003 г., 1 сентября; 4(5):421-5. doi: 10.1046/j.1364-3703.2003.00179.x. Мол Плант Патол. 2003. PMID: 20569401
Дифференциальная индукция TNF-альфа и IGTP, IIGP структурно разнообразными классическими и неклассическими липополисахаридами.
Лапак Н., Такеучи О., Корралес Ф., Акира С., Морион И., Ховард Дж. К., Горвел Дж. П. Лапак Н. и др. Клеточная микробиология. 2006 март;8(3):401-13. doi: 10.1111/j.1462-5822.2005.00629.x. Клеточная микробиология. 2006. PMID: 16469053
Разнообразие эндотоксинов и их влияние на патогенез.
Trent MS, Stead CM, Tran AX, Hankins JV. Трент М.С. и др. J Эндотоксин Res. 2006;12(4):205-23.
дои: 10.1179/096805106X118825.
J Эндотоксин Res. 2006.
PMID: 16953973
Обзор.Разнообразие основного олигосахарида в липополисахаридах.
Силипо А., Молинаро А. Силипо А и др. Субклеточная биохимия. 2010;53:69-99. doi: 10.1007/978-90-481-9078-2_4. Субклеточная биохимия. 2010. PMID: 20593263
Посмотреть все похожие статьи
Цитируется
Xanthomonas hortorum — за пределами садов: текущая таксономия, геномика и репертуар вирулентности.
Диа Н.К., Мориньер Л., Коттин Б., Берналь Э., Джейкобс Дж. М., Коебник Р., Осдаги Э., Потнис Н., Потье Дж. Ф. Диа Н.К. и др. Мол Плант Патол. 2022 май; 23(5):597-621. doi: 10.1111/mpp.13185. Epub 2022 23 января.
Мол Плант Патол. 2022.
PMID: 35068051
Бесплатная статья ЧВК.Полногеномная идентификация генов Ralstonia solanacearum, необходимых для выживания растений томатов.
Су И, Сюй И, Лян Х, Юань Г, У С, Чжэн Д. Су Ю и др. mSystems. 26 октября 2021 г.; 6(5):e0083821. doi: 10.1128/mSystems.00838-21. Epub 2021 12 октября. mSystems. 2021. PMID: 34636662 Бесплатная статья ЧВК.
Сравнительное исследование химического состава и биологической активности сосны ( Pinus nigra и P. sylvestris ) Экстракты коры.
Нишка А., Штефэнеску Р., Стегэруш Д.И., Маре А.Д., Фарчади Л., Тэнасе К. Ниска А. и др. Антиоксиданты (Базель). 2021 22 февраля; 10 (2): 327. doi: 10.3390/antiox10020327. Антиоксиданты (Базель).
2021.
PMID: 33671773
Бесплатная статья ЧВК.Геномное разнообразие и организация сложных кластеров биосинтеза полисахаридов у рода Dickeya.
Ранджан М., Хохани Д., Наяка С., Шривастава С., Кейсер З.П., Ранджан А. Ранджан М. и др. ПЛОС Один. 2021 11 февраля; 16 (2): e0245727. doi: 10.1371/journal.pone.0245727. Электронная коллекция 2021. ПЛОС Один. 2021. PMID: 33571209 Бесплатная статья ЧВК.
Липополисахарид усиливает биосинтез таншинона посредством Ca 2+ -зависимого способа в волосистых корнях Salvia miltiorrhiza .
Чжан Б., Ли Х, Ли Х, Лу Зи, Цай Х, Оу Ян К, Ма П, Донг Дж. Чжан Б. и др. Int J Mol Sci. 2020 16 декабря; 21 (24): 9576. дои: 10.3390/ijms21249576. Int J Mol Sci.