
Пластинчатый теплообменник или трубчатый. Что выбрать?
Не более чем, 20 лет назад вопрос: что выбрать трубчатый теплообменник или пластинчатый, не существовал. В большинстве случаев применялись трубчатые теплообменники. Но ничего не стоит на месте, со временем появилось более эффективное теплообменное оборудование. На смену трубчатым теплообменникам стали использовать пластинчатые. В этой статье мы постараемся разобраться в преимуществах и недостатках обоих видов теплообменников.
Помогать нам будет инженер – теплотехник Белгородских тепловых сетей, Сизов Петр Михайлович. Он любезно ответил на наши вопросы, и мы надеемся, эти ответы помогут нашим заказчикам с выбором необходимого им теплообменного оборудования.
Как происходит процесс теплообмена в трубчатом теплообменнике?
— Процесс теплообмена в таких теплообменниках происходит через теплопередающую поверхность (поверхность трубок внутри теплообменника).
Рассмотрим пример нагрева. Греющий теплоноситель от источника тепла поступает в межтрубное пространство, в котором расположены трубки (трубное пространство). По этим трубкам протекает нагреваемый теплоноситель. Греющий теплоноситель проходит по межтрубному пространству и контактирует с поверхностью трубок, таким образом он он нагревает вторую среду. Среды в таком теплообменнике движутся в противотоке.
Как работает пластинчатый теплообменник?
— В пластинчатом теплообменнике среды обмениваются теплом тоже через теплопередающую поверхность, которой являются пластины теплообменника. Пластины при сборке образуют пакет, в котором есть каналы. По этим каналам протекают греющий и нагреваемый теплоносители.
Рассмотрим пример нагрева. Условно, возьмем для примера одну пластину. С одной стороны этой пластины по каналам будет протекать греющий теплоноситель.
Какие сильные и слабые стороны у трубчатых теплообменников?
Преимущества:
— Хорошая стойкость к гидроударам. Высокие рабочие характеристики по давлению.
— Небольшая разница температур греющего и нагреваемого теплоносителя (1С — 2С)
— Менее требовательны к качеству воды. Но это пункт спорный. Т.к. забитый теплообменник резко теряет свои теплопередающие свойства.
Недостатки:
— Низкий коэффициент теплопередачи. Эффективность нагрева хуже, чем у пластинчатых теплообменников.
— Большие габаритные размеры. Большой вес и металлоемкость конструкции. Требует большой площади для монтажа и специального фундамента.
— При ремонте, не редко, проводят заглушку вышедших из строя трубок. Это снижает теплопередающую поверхность теплообменника.
Это снижает теплопередающую поверхность теплообменника.
— Течь теплообменника в трубном пространстве можно обнаружить только при разборке теплообменника
— Промывку можно проводить только безразборным способом. Наиболее сложно удалять отложения в трубках теплообменника, особенно когда они сильно зарастают накипью.
— Необходима теплоизоляция конструкции
— Невозможно увеличить поверхность теплопередачи.
Какие преимущества и недостатки у пластинчатых теплообменников?
Преимущества:
— КПД 80-85%.
— Коэффициент теплопередачи выше чем у трубчатых теплообменник. Эффективность нагрева лучше.
— Компактные размеры и патрубки, расположенные с одной стороны. Это экономит место и упрощает монтаж оборудования.
— Есть возможность увеличить мощность теплообменника. Для этого необходимо, просто, добавить пластины.
— Простота сервисного обслуживания. Промывку теплообменника можно проводить разборным способом, силами 2 человек.
— Не требуют теплоизоляции.
— Пониженная загрязняемость. Среды при движении в каналах достигают высокой степени турбулентности. Это уменьшает образование отложений на поверхности пластин.
Недостатки:
— Относительно высокие гидравлические потери.
— Более требовательны к качеству воды, чем трубчатые.
—
Какое оборудование дешевле?
При сравнении оборудования работающего для систем горячего водоснабжения и отопления, т.е. при использовании в системах ЖКХ — трубчатые теплообменники будут незначительно дешевле пластинчатых теплообменников. Но сравнение только закупочной цены – это неверный метод. Необходимо сравнивать цену и стоимость сервисного обслуживания оборудования.
Если сравнивать по схеме: закупочная цена + цена эксплуатации оборудования, то пластинчатые теплообменники, явно выигрывают.
Какие теплообменники проще обслуживать?
В силу своей разборной конструкции, пластинчатые теплообменники проще обслуживать. А эффективность промывки выше, потому что при разборной промывке мы можем визуально проконтролировать качество проведенных работ. Все детали в разборных теплообменниках легко заменить при ремонте.
А эффективность промывки выше, потому что при разборной промывке мы можем визуально проконтролировать качество проведенных работ. Все детали в разборных теплообменниках легко заменить при ремонте.
Какой срок эксплуатации?
Правильнее будет использовать термин «межремонтный период», т.е. срок до проведения капитального ремонта. По заявлениям производителей, у трубчатых теплообменников этот период 5-10 лет. У пластинчатых 10-15 лет.
Какие выводы необходимо сделать?
На данный момент идут разработки трубчатых теплообменников, которые по своим характеристикам приближены к пластинчатым. Но цена и стоимость обслуживания такого оборудования пока не известны. Поэтому, для работы в системах ЖКХ, паровых системах, промышленности наилучшим выбором будет разборный пластинчатый теплообменник.
Если ваше оборудование будет работать в условиях повышенного давления (более 25 бар) или повышенной температуры (более 200С), то необходимо обратить внимание на трубчатые теплообменники.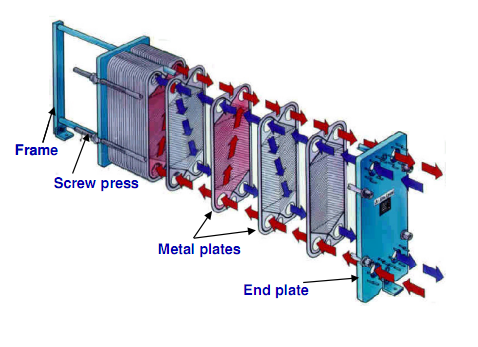
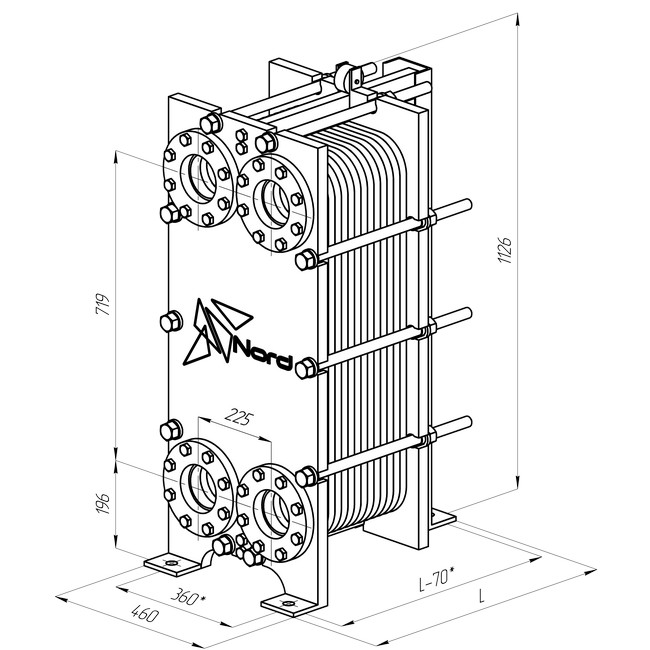
Пластинчатые теплообменники — Пластинчатый теплообменник TG077P
Пластинчатый теплообменник TG077P
Количество пластин
21 23 25 27 29 31 33 35 37 41 45 49 51 55 61 65 71 75 81 85 91 95 101
Присоединения
Штуцер AISI304 1″ Штуцер AISI304 1¼» Штуцер AISI304 1½»
Площадь пластины 0,07 м2
Материал пластин: Нерж. сталь AISI316
сталь AISI316
Толщина пластин: 0,4мм, 0,5мм
Материал уплотнений: EPDM, NBR
Присоединения: Наружная резьба 1″, 1¼», 1½»
Материал штуцеров: Нерж.сталь AISI304
Площадь пластины 0,07 м2
Материал пластин: Нерж.сталь AISI316
Толщина пластин: 0,4мм, 0,5мм
Материал уплотнений: EPDM, NBR
Присоединения: Наружная резьба 1″, 1¼», 1½»
Материал штуцеров: Нерж.сталь AISI304
Пластинчатый теплообменник Danfoss D55-H50 (021h4625)
Теплообменники Danfoss MPHE отличаются более интенсивным потоком, который проходит через пластину, и оптимальным использованием всей её поверхности. Это существенно улучшает теплопередачу. По сравнению с традиционными паяными пластинчатыми теплообменниками, показатель эффективности теплообменника (EI) у теплообменников MPHE, при одинаковых условиях работы, намного выше.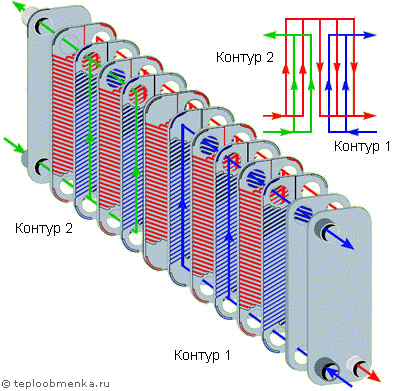 В некоторых случаях это преимущество достигает 56%.
В некоторых случаях это преимущество достигает 56%.
Меньшее количество используемого хладагента, улучшенная теплопередача, более экономное использование материалов, отрегулированные перепады давления – все это делает новую технологию компании «Данфосс» отличным выбором для тех, кто стремится к получению повышенной производительности при соблюдении экологической безопасности.
ПРЕИМУЩЕСТВА ТЕПЛООБМЕННИКОВ Danfoss MPHE:
– Уникальное расположение каналов теплообменников MPHE, производимых компанией «Данфосс», позволяет расширить границы существующих технологий. Теплообменники Danfoss демонстрируют повышенную эффективность работы при минимизированных затратах ресурсов, а также отличаются повышенной теплопередачей. Таким образом, Вы приближаетесь к желаемой цели – разумному использованию ресурсов.
– Теплообменники MPHE предоставляют отличные возможности для реализации различных конструкторских решений. Всего лишь меняя количество, размер и расположение каналов на гофрированной пластине, данные теплообменники можно модифицировать таким образом, что они будут иметь самые высокие показатели теплопередачи и минимальные перепады давления при любых вариантах использования. Первые модели, которые компания «Данфосс» предлагает на рынке – серия «Н», являющаяся оптимальным техническим решением для тепловых насосов и серия «С», являющаяся идеальным вариантом для использования в водоохладителях. Разрабатываются также и другие модели.
Всего лишь меняя количество, размер и расположение каналов на гофрированной пластине, данные теплообменники можно модифицировать таким образом, что они будут иметь самые высокие показатели теплопередачи и минимальные перепады давления при любых вариантах использования. Первые модели, которые компания «Данфосс» предлагает на рынке – серия «Н», являющаяся оптимальным техническим решением для тепловых насосов и серия «С», являющаяся идеальным вариантом для использования в водоохладителях. Разрабатываются также и другие модели.
– Компания Danfoss предлагаеn серию различных теплообменников MPHE, предназначенных для тепловых насосов, водоохладителей и множества других применений. И всегда рада помочь Вам выбрать нужную модель или изменить дизайн гофрированной пластины, чтобы добиться оптимальной работы производимого Вами оборудования.
– Теплообменники компании Danfoss MPHE значительно компактнее кожухотрубных теплообменников и надёжнее паяных пластинчатых теплообменников (отсутствуют уплотнения, которые могут выйти из строя). Теплообменники MPHE способны работать при более высоких температурах и давлениях.
Теплообменники MPHE способны работать при более высоких температурах и давлениях.
– Поскольку теплообменники MPHE разработаны на основе испытанной и проверенной временем технологии паяных пластин, они действительно надежны, экономичны и эффективны. Теплообменники MPHE отличаются от паяных пластинчатых теплообменников специфичным расположением каналов в пластинах, что даёт улучшенный теплообмен и делает конструкцию более гибкой для нахождения оптимальных решений в различных применениях.
– По сравнению с паяными пластинчатыми теплообменниками, теплообменники MPHE имеют меньший внутренний объем и меньшее количество холодильного агента в системе. Поэтому они быстрее реагируют на изменения температуры и имеют меньший углеродный след.
– Вы можете не сомневаться в том, что компания Danfoss предлагает изделия наивысшего качества, поскольку, перед поставкой заказчику, все теплообменники MPHE проходят испытания давлением на прочность и на отсутствие утечек. Теплообменники Danfoss соответствуют всем распространенным нормам для сосудов, работающих под давлением, включая такие, как Директива ЕС для оборудования, работающего под давлением, стандартам UL/CSA и КНК.
Теплообменники Danfoss соответствуют всем распространенным нормам для сосудов, работающих под давлением, включая такие, как Директива ЕС для оборудования, работающего под давлением, стандартам UL/CSA и КНК.
– Заказчики компании Danfoss могут чувствовать себя спокойно, поскольку теплообменники MPHE соответствуют экологическим нормам и стандартам качества ISO 14000 и ISO 9001.
- Видео
- Каталоги
- Технические характеристики
- Размеры и подключения
| Модель | D55-H50 |
| Код | 021h4625 |
| Количество пластин | 50 |
| Мощность, кВ (испаритель) | – |
| Мощность, кВ (конденсатор) | 25,72 |
Высота, мм. | 525 |
| Ширина, мм. | 109 |
| Вес, кг. | 10,9 |
| Комб. партубок (пайка) Q3 – хол. вх. | ISO-G1 / 7/8″ |
| Комб. партубок (пайка) Q4 – хол. вых. | ISO-G1 / 7/8″ |
| Комб. партубок (резьба/пайка) Q1 | ISO-G1 / 7/8″ |
| Комб. партубок (резьба/пайка) Q2 | ISO-G1 / 7/8″ |
| Минимальная рабочая температура, ºС | -196 |
| Максимальная рабочая температура, ºС | 200 |
| Максимальное рабочее давление, бар | 30 |
Рассчетные условия (конденсатор):
- Сторона 1: R404a; Tконд. = 40ºС; Перегрев = 2K
- Сторона 2: Tвх. = 32 ºС; Tвых. = 37 ºС; ΔP<40 kPa
Теплообменники D55 поставляются в различных конфигурациях:
- D55: Конфигурация, сертифицированная на давление до 30 бар.
- D55-E: Испаритель, сертифицированный на давление до 30 Бар, с распределителем хладагента.

- D55-H: Конфигурация, сертифицированная на давление до 30 Бар, со схемой расположения каналов «high-theta».
- D55L: Конфигурация, сертифицированная на давление до 45 бар.
- D55L-E: Испаритель, сертифицированный на давление до 45 Бар, с распределителем хладагента.
- D55L-H: Конфигурация, сертифицированная на давление до 45 Бар, со схемой расположения каналов «high-theta».
| Модель | A(мм/дюймы) | Мин./Макс. Рабочая температура | Макс. рабочее давление | Поддерживаемый объём: Q1-Q2 / Q3-Q4 | Вес: |
| D55 | 9,5+1.74xn / 0,37+0.07xn | -196 °C/-320 °F | 30 bar/435 psi | (l): 0,076хn/2 / 0,076х(n-2)/2 (ft3): 0,003хn/2 / 0,003х(n-2)/2 | 2,32кг+0,135xn / 5,12фунт+0,30xn |
| D55-E | |||||
| D55-H | 9,5+1,44xn / 0,37+0,06xn | (l): 0,061хn/2 / 0,061х(n-2)/2 (ft3): 0,002хn/2 / 0,002х(n-2)/2 | |||
| D55-L | 11,5+1. 74xn / 0,45+0.07xn 74xn / 0,45+0.07xn | 45 bar/650 psi | (l): 0,076хn/2 / 0,076х(n-2)/2 (ft3): 0,003хn/2 / 0,003х(n-2)/2 | 2,67кг+0.135xn / 5,89кг+0.30xn | |
| D55L-E | |||||
| D55L-H | 11,5+1.44n / 0,45+0.06n | (l): 0,061хn/2 / 0,061х(n-2)/2 (ft3): 0,002хn/2 / 0,002х(n-2)/2 | |||
n = число пластин
Пластинчатый теплообменник Danfoss D55-H50 (021h4625)
[contact-form-7 404 «Not Found»]ГК ТЕПЛОСИЛА ET-002 DN25 Теплообменник пластинчатый
Теплообменник пластинчатый разборный ET-002 DN25 предназначен для осуществления процесса теплообмена между жидкими средами в системах отопления, горячего водоснабжения и вентиляции жилых, административных и промышленных зданий, а также в различных технологических теплообменных процессах. Теплообменник данного типа не предназначен для работы с токсичными, взрывоопасными и пожароопасными средами.
Храктеристика | Значение |
|---|---|
Максимальное количество пластин, шт. | 160 |
Максимальная площадь теплообмена, m2 | 4,3 |
Максимальный расход, m3/ч | 5 |
Условный диаметр патрубка, мм | 25 |
Вес, кг, не более | 43 |
Рабочее давление, бар (МПа) | 16 (1,6) |
Температура рабочей среды, °C | -10…+150 |
Рабочие среды | вода, этиленгликоль и пропиленгликоль (концентрация до 65%) |
Материал резиновых уплотнений | резина марки EPDM |
Материал пластин | нержавеющая сталь AISI 304, AISI 316 |
Россия, Казахстан, Белоруссия, Узбекистан, Армения, Киргизия, Таджикистан — доставка в любой город и другие страны ЕАЭС и мира.
Имя должно быть не менее :error символов.
Не правильный E-mail.
Название должно быть не менее :error символов.
Обязательное поле
Защита от спама reCAPTCHA Конфиденциальность и Условия использования
Сообщение отправлено
Пожалуйста, заполните форму правильно.
Отправка…
Капча недействительна.
Повторите попытку позже.
ОПЫТ
ГК Теплосила имеет более чем 20-летний опыт в сфере производства оборудования для тепловых пунктов. Продукция эффективно применяется в России, Беларуси, Казахстане, Монголии и других странах.
КАЧЕСТВО
Продукция Теплосила соответствует международным стандартам качества, она проходит несколько этапов технического контроля, тестирование в собственной лаборатории и сертификацию в соответствии с требованиями ГОСТ.
СЕРВИС
ГК Теплосила обеспечивает своим клиентам первоклассный сервис, профессиональное обслуживание и техническую поддержку. Благодаря этому оборудование работает эффективно и исправно.
Теплообменники пластинчатые
ET-002, ET-010, ET-015M и др.
Регуляторы перепада давления
RDT, RDT-S, RDT-P и др.

Регулирующие клапаны
TRV, TRV-3 и др.
Модули управления
TTR-01D, TTR-01A и др.
Шкафы управления
ТШУ-1-1, ТШУ-1-2 и др.
Имя должно быть не менее :error символов.
Не правильный E-mail.
Название должно быть не менее :error символов.
Обязательное поле
Защита от спама reCAPTCHA Конфиденциальность и Условия использования
Сообщение отправлено
Пожалуйста, заполните форму правильно.
Отправка…
Капча недействительна.
Повторите попытку позже.
адрес для заявок: [email protected]- (7273)495-231
- (3955)60-70-56
- (8182)63-90-72
- (8512)99-46-04
- (3852)73-04-60
- (4722)40-23-64
- (4162)22-76-07
- (4832)59-03-52
- (423)249-28-31
- (8672)28-90-48
- (4922)49-43-18
- (844)278-03-48
- (8172)26-41-59
- (473)204-51-73
- (343)384-55-89
- (4932)77-34-06
- (3412)26-03-58
- (395)279-98-46
- (843)206-01-48
- (4012)72-03-81
- (4842)92-23-67
- (3842)65-04-62
- (8332)68-02-04
- (4966)23-41-49
- (4942)77-07-48
- (861)203-40-90
- (391)204-63-61
- (4712)77-13-04
- (3522)50-90-47
- (4742)52-20-81
- (3519)55-03-13
- (495)268-04-70
- (8152)59-64-93
- (8552)20-53-41
- (831)429-08-12
- (3843)20-46-81
- (3496)41-32-12
- (383)227-86-73
- (3812)21-46-40
- (4862)44-53-42
- (3532)37-68-04
- (8412)22-31-16
- (8142)55-98-37
- (8112)59-10-37
- (342)205-81-47
- (863)308-18-15
- (4912)46-61-64
- (846)206-03-16
- (8342)22-96-24
- (812)309-46-40
- (845)249-38-78
- (8692)22-31-93
- (3652)67-13-56
- (4812)29-41-54
- (862)225-72-31
- (8652)20-65-13
- (3462)77-98-35
- (8212)25-95-17
- (4752)50-40-97
- (4822)63-31-35
- (8482)63-91-07
- (3822)98-41-53
- (4872)33-79-87
- (3452)66-21-18
- (8422)24-23-59
- (3012)59-97-51
- (347)229-48-12
- (4212)92-98-04
- (8352)28-53-07
- (351)202-03-61
- (8202)49-02-64
- (3022)38-34-83
- (4112)23-90-97
- (4852)69-52-93
Задайте вопрос прямо сейчас:
Извините, сервис временно недоступен.
Некорректный номер.
Ожидайте звонка на введенный номер.
front/header.call_free_error
Заказать обратный звонок
мы перезвоним Вам в рабочее время
Настоящее соглашение является официальным документом OOO «Новые Технологии», ОГРН 1131690023178, ИНН 1656069657 (далее – Администратор) и определяют порядок использования посетителями (далее — Посетитель) сайта Администратора и обработки информации, получаемой Администратором от Посетителя.
- Соглашение может быть изменено Администратором в одностороннем порядке в любой момент, без какого-либо специального уведомления Посетителя Сайта.
- В случае, если при использовании Посетителями Сайта Администратору будет сообщена какая-либо информация, относящаяся прямо или косвенно к определенному или определяемому физическому лицу (далее – Персональные данные), ее последующая обработка будет осуществляться в соответствии с законодательством Российской Федерации.
 В отношении всех сообщаемых Персональных данных Посетитель дает Администратору согласие на их обработку. Администратор обрабатывает персональные данные Посетителя исключительно в целях предоставления Посетителю функций Сайта, размещенного на нем контента, маркетинговой, рекламной, иной информации, в целях получения Посетителем персонализированной (таргетированной) рекламы, исследования и анализа данных Посетителя, а также в целях предложения Посетителю своих товаров и услуг. В отношении всех сообщенных Администратору Посетителем своих персональных данных Администратор вправе осуществлять сбор, систематизацию, накопление, хранение, уточнение (обновление, изменение), использование, распространение (в том числе передача любым третьим лицам, включая передачу персональных данных третьим лицам на хранение или в случае поручения обработки персональных данных третьим лицам), обезличивание, блокирование, уничтожение, трансграничную передачу, обработку с применением основных способов такой обработки (хранение, запись на электронных носителях и их хранение, составление перечней, маркировка) и иные действия в соответствии со статьей 3 Федерального закона от 27.
В отношении всех сообщаемых Персональных данных Посетитель дает Администратору согласие на их обработку. Администратор обрабатывает персональные данные Посетителя исключительно в целях предоставления Посетителю функций Сайта, размещенного на нем контента, маркетинговой, рекламной, иной информации, в целях получения Посетителем персонализированной (таргетированной) рекламы, исследования и анализа данных Посетителя, а также в целях предложения Посетителю своих товаров и услуг. В отношении всех сообщенных Администратору Посетителем своих персональных данных Администратор вправе осуществлять сбор, систематизацию, накопление, хранение, уточнение (обновление, изменение), использование, распространение (в том числе передача любым третьим лицам, включая передачу персональных данных третьим лицам на хранение или в случае поручения обработки персональных данных третьим лицам), обезличивание, блокирование, уничтожение, трансграничную передачу, обработку с применением основных способов такой обработки (хранение, запись на электронных носителях и их хранение, составление перечней, маркировка) и иные действия в соответствии со статьей 3 Федерального закона от 27. 07.2006 № 152-ФЗ «О персональных данных».
07.2006 № 152-ФЗ «О персональных данных». - Посетитель понимает и соглашается с тем, что предоставление Администратору какой-либо информации о себе, не являющейся контактной и не относящейся к целям, обозначенным Администратором Сайта (не относящейся к деятельности Администратора, к продвигаемым им товарам и/или услугам, к условиям сотрудничества Администратора и Посетителя Сайта), а равно предоставление информации, относящейся к государственной, банковской и/или коммерческой тайне, информации о расовой и/или национальной принадлежности, политических взглядах, религиозных или философских убеждениях, состоянии здоровья, интимной жизни Посетителя Сайта или иного третьего лица запрещено.
- В случае принятия Посетителем решения о предоставлении Администратору какой-либо информации (каких-либо данных), Посетитель обязуется предоставлять исключительно достоверную и актуальную информацию. Посетитель Сайта не вправе вводить Администратора в заблуждение в отношении своей личности, сообщать ложную или недостоверную информацию о себе.

- Администратор принимает меры для защиты Персональных данных Посетителя Сайта в соответствии с законодательством Российской Федерации.
- Администратор не проверяет достоверность персональной информации, предоставляемой Посетителем Сайта, и не имеет возможности оценивать его дееспособность. Однако Администратор исходит из того, что Посетитель предоставляет достоверную персональную информацию и поддерживает эту информацию в актуальном состоянии.
- Администратор вправе запрещать Посетителю доступ к Сайту или к отдельным частям Сайта.
- Посетитель в соответствии с ч. 1 ст. 18 Федерального закона «О рекламе» дает Администратору свое согласие на получение сообщений рекламного характера.
- Согласие может быть отозвано субъектом персональных данных или его представителем путем направления письменного заявления ООО «Новые Технологии» или его представителю по адресу: 420030 Казань, Адмиралтейская д.
 3 к.4 п.1026.
3 к.4 п.1026. - В случае отзыва субъектом персональных данных или его представителем Согласия на обработку персональных данных, ООО «Новые Технологии» вправе продолжить обработку без разрешения субъекта персональных данных при наличии оснований, указанных в пунктах 2 — 11 части 1 статьи 6, части 2 статьи 10 и части 2 статьи 11 Федерального закона №152-ФЗ «О персональных данных» от 26.06.2006 г.
- Настоящее Согласие действует все время до момента прекращения обработки персональных данных по причинам, указанным в п. 9 данного документа.
Разборные пластинчатые теплообменники Alfa Laval — каталог, технические характеристики, запрос КП
Разборные пластинчатые теплообменники Alfa Laval — каталог, технические характеристики, запрос КП | Alfa Laval- Оборудование
- Сервис
- Отрасли промышленности
- Принцип работы
- Сертификаты и брошюры
- Видео
- Проверка герметичности
Разборные пластинчатые теплообменники обеспечивают эффективную передачу тепла, обладая при этом компактными размерами и малой занимаемой площадью. Эти устройства имеют легко приспосабливаемую под нужды процесса гибкую конструкцию и не требуют сложного технического обслуживания. Представленные в чрезвычайно широком ассортименте, они применяются для нагрева, охлаждения, утилизации тепла, испарения и конденсации во множестве областей.
Эти устройства имеют легко приспосабливаемую под нужды процесса гибкую конструкцию и не требуют сложного технического обслуживания. Представленные в чрезвычайно широком ассортименте, они применяются для нагрева, охлаждения, утилизации тепла, испарения и конденсации во множестве областей.
Преимущества
- Высочайшая тепловая эффективность и малая разница температур
- Компактный размер — экономия места и простота и обслуживания
- Максимальное время безотказной работы — меньше загрязнений, напряжений, износа и коррозии
- Гибкость — легко адаптируется к изменившимся техническим условиям
- Большинство моделей производится на заводе в России
Наши разборные пластинчатые теплообменники оптимизируют теплопередачу благодаря обширным поверхностям профилированных пластин, передающих тепло от одних газов или жидкостей к другим. Мы предлагаем высокую эффективность, непревзойденную надежность на протяжении всего срока службы, а также компактное исполнение по разумной цене. Вы всегда можете заказать конструкцию, которая легко адаптируется под нужды вашего процесса. Также среди преимуществ наших теплообменников: энергосберегающие свойства и низкий углеродный след.
Вы всегда можете заказать конструкцию, которая легко адаптируется под нужды вашего процесса. Также среди преимуществ наших теплообменников: энергосберегающие свойства и низкий углеродный след.
Промышленная серия
Модельный ряд промышленной серии разборных пластинчатых теплообменников Alfa Laval используется для различных применений для решения таких задач, как нагрев, охлаждение, рекуперация тепла, конденсация и испарение.
Модельный ряд
Новые стандарты для ваших теплообменников
Представляем самые современные в мире разборные пластинчатые теплообменники. Узнайте, как наша линейка разборных пластинчатых теплообменников следующего поколения обеспечит более высокую эффективность, повышенную надежность и большее удобство обслуживания.
Подробнее
Конструкция пластин с профилированной поверхностью оптимизирует теплопередачу, обеспечивая большую, но при этом компактную общую площадь поверхности, через которую тепло может передаваться от одной жидкости/газа к другой/другому.
Область теплопередачи пластин спрессована «елочкой». При наложении двух пластин друг на друга с противоположно направленным рисунком-«елочкой» спиральный поток и высокая турбулентность обеспечивают высокие коэффициенты переноса и эффективную самоочистку.
Изменение рисунка позволяет использовать теплообменник в различных процессах, даже с очень грязной средой.
Распределительная площадка
Распределительная площадка на пластине обеспечивает равномерный поток жидкости по всем пластинам, оптимизируя теплопередачу. Оптимизированное распределение потока предотвращает загрязнение и как следствие образование участков с различной температурой, поддерживая высокий уровень производительности на протяжении всего срока службы без энергопотерь, расходов на техническое обслуживание и незапланированных остановок.
Уплотнения — это ключевые компоненты для производительности теплообменников. Мы конструируем уплотнение и пластины как единое целое, чтобы гарантировать оптимизированную изоляцию. Все наши решения разработаны с учетом назначения теплообменника. Правильный профиль, ширина, толщина, а также тип полимера и компаунд существенно снижают риск преждевременной утечки или повреждения уплотнения или пластин. Наши решения гарантируют вам увеличенный срок службы уплотнения, меньшее время простоя и снижение расходов на техническое обслуживание.
Специальные типы пластин
Пластины с двойной стенкой для разборных теплообменников
Разработаны для использования с жидкостями, которые не должны смешаться даже в случае протечки. Предотвращают контакт просачивающейся жидкости с жидкостью в другом контуре.
Пластины для теплообменников с большим каналом между пластинами Wide-gap
Идеально для жидкостей, содержащих волокна или жесткие частицы. Разработаны для предотвращения скопления твердых веществ в области входа.
Разработаны для предотвращения скопления твердых веществ в области входа.
Узнайте больше о теплообменниках WideGap
Пластины для полусварных теплообменников
Позволяют работать с агрессивными и сложными жидкостями, а также увеличивают диапазон давления.
Узнайте больше о полусварных теплообменниках
Пластины из графитового материала для теплообменников Diabon®
Композит из плавленого графита и фторсодержащего полимера обеспечивает отличную стойкость к коррозионным материалам.
Узнайте больше о пластинах из графитового материала для теплообменников Diabon
.
Концепция теплообменника относительно проста — нагрев или охлаждение одной среды путем передачи тепла между ней и еще одной средой.
В разборных пластинчатых теплообменниках пластины снабжены эластомерными уплотнениями, которые запечатывают каналы и направляют среду в альтернативные каналы. Пакеты пластин смонтированы между опорной и нажимной плитами и стягиваются стяжными болтами, закрепленными между этими плитами. Каналы, образованные пластинами, и прижимные плиты размещены на верхней несущей балке и зафиксированы на поддерживающей колонне. Конструкция позволяет легко проводить чистку и изменять площадь (добавляя или удаляя пластины).
Каналы, образованные пластинами, и прижимные плиты размещены на верхней несущей балке и зафиксированы на поддерживающей колонне. Конструкция позволяет легко проводить чистку и изменять площадь (добавляя или удаляя пластины).
Зона теплопередачи разборного пластинчатого теплообменника состоит из ряда пластин с профилированной поверхностью, установленных между рамой и прижимными пластинами для удержания давления. Уплотнения действуют как герметизирующие прокладки между пластинами. Жидкости обычно проходят через теплообменник во встречном направлении. Это обеспечивает максимальную теплоэффективность и маленькую температурную дельту между первичной средой на выходе и вторичной средой на входе.
Для термочувствительных или вязких сред может использоваться сонаправленные потоки, чтобы самая холодная среда могла встретиться с самой горячей на входе в теплообменник. Это минимизирует риск перегрева или обмерзания.
Пластины предлагаются с разными вариантами глубинами штамповки, углом и шевронами, что позволяет обеспечить оптимальную производительность. Для каждой линейки продуктов, в зависимости от применения, пластины обладают собственными преимуществами.
Для каждой линейки продуктов, в зависимости от применения, пластины обладают собственными преимуществами.
Распределительная площадка обеспечивает равномерное распределение жидкостей по всей поверхности теплопередачи и помогает избежать образования зон застаивания, способных спровоцировать загрязнение.
Высокая турбулентность потока между пластинами обусловливает высокую степень теплообмена, что в свою очередь приводит к перепадам давления. Наши инженеры по тепловым расчетам помогут спроектировать и выбрать ту модель и конфигурацию, которые подойдут к вашему применению и обеспечат максимальную теплоэффективность при минимальном перепаде давления.
Стандартные разборные пластинчатые теплообменники
Гигиенические применения
Специальные применения
Сделай это вовремя. Проверка герметичности пластин Вашего теплообменника.

Разборные пластинчатые теплообменники надежны, но, как и любое оборудование, требуют надлежащего технического обслуживания. Проверка герметичности от Альфа Лаваль поможет вам выявить потенциальные проблемы, такие как трещины или утечки, чтобы вы могли вовремя принять необходимые меры и избежать неожиданных поломок.
Узнайте больше
.
Теплообменник пластинчатый Aquaviva, AISI 316L
Технические характеристики
пластинчатых теплообменников Aquaviva, AISI 316L
| Модель | 140 кВт | 242 кВт | 287 кВт | 446 кВт | 672 кВт | 819 кВт |
|---|---|---|---|---|---|---|
| Мощность, кВт | 140 | 242 | 287 | 446 | 672 | 819 |
| Площадь теплопередачи, м² | 1. 13 13 |
0.95 | 1.1 | 1.68 | 2.37 | 2.99 |
| Материал пластин | нержавеющая сталь AISI 316L | нержавеющая сталь AISI 316L | нержавеющая сталь AISI 316L | нержавеющая сталь AISI 316L | нержавеющая сталь AISI 316L | нержавеющая сталь AISI 316L |
| Количество пластин, шт | 29 | 29 | 29 | 29 | 29 | 29 |
| Толщина пластин, мм | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Вес пустого теплообменника, кг | 33.19 | 63.93 | 64.9 | 68.78 | 113.93 | 118.22 |
| Вес полного теплообменника, кг | 37.85 | 66.5 | 67.87 | 73.33 | 120. 83 83 |
126.92 |
| Объем контура горячей/холодной стороны, л | 4.66 | 2.57 | 2.96 | 4.54 | 6.9 | 8.7 |
| Рабочая температура, °C | от -20 до +109 | от -20 до +109 | от -20 до +109 | от -20 до +109 | от -20 до +109 | от -20 до +109 |
| Рабочее давление, бар | 10 | 10 | 10 | 10 | 10 | 10 |
| Тестовое давление, бар | 14.3 | 14.3 | 14.3 | 14.3 | 14.3 | 14.3 |
| Горячая сторона | ||||||
| Температура воды на входе/выходе, °C | 90/70 | 90/70 | 90/70 | 90/70 | 90/70 | 90/70 |
| Скорость протока, м³/ч | 6. 17 17 |
10.67 | 12.65 | 19.66 | 29.63 | 36.11 |
| Перепад давления, бар | 0.23 | 0.21 | 0.23 | 0.26 | 0.27 | 0.28 |
| Холодная сторона | ||||||
| Температура воды на входе/выходе, °C | 20/35 | 20/35 | 20/35 | 20/35 | 20/35 | 20/35 |
| Скорость протока, м³/ч | 8.07 | 13.95 | 13.55 | 25.71 | 38.74 | 47.22 |
| Перепад давления, бар | 0.49 | 0.41 | 0.44 | 0.49 | 0.48 | 0.49 |
Свернуть таблицу Развернуть таблицу
| Модель | 140 кВт |
|---|---|
| Мощность, кВт | 140 |
| Площадь теплопередачи, м² | 1. 13 13 |
| Материал пластин | нержавеющая сталь AISI 316L |
| Количество пластин, шт | 29 |
| Толщина пластин, мм | 0.5 |
| Вес пустого теплообменника, кг | 33.19 |
| Вес полного теплообменника, кг | 37.85 |
| Объем контура горячей/холодной стороны, л | 4.66 |
| Рабочая температура, °C | от -20 до +109 |
| Рабочее давление, бар | 10 |
| Тестовое давление, бар | 14.3 |
| Горячая сторона | |
| Температура воды на входе/выходе, °C | 90/70 |
| Скорость протока, м³/ч | 6.17 |
| Перепад давления, бар | 0.23 |
| Холодная сторона | |
| Температура воды на входе/выходе, °C | 20/35 |
| Скорость протока, м³/ч | 8.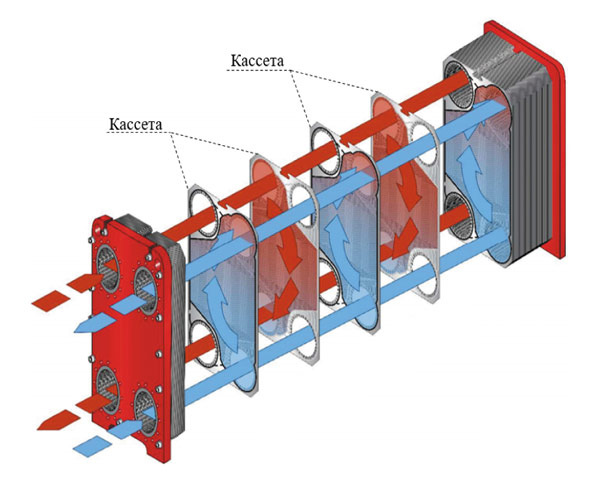 07 07 |
| Перепад давления, бар | 0.49 |
| Модель | 242 кВт |
|---|---|
| Мощность, кВт | 242 |
| Площадь теплопередачи, м² | 0.95 |
| Материал пластин | нержавеющая сталь AISI 316L |
| Количество пластин, шт | 29 |
| Толщина пластин, мм | 0.5 |
| Вес пустого теплообменника, кг | 63.93 |
| Вес полного теплообменника, кг | 66.5 |
| Объем контура горячей/холодной стороны, л | 2.57 |
| Рабочая температура, °C | от -20 до +109 |
| Рабочее давление, бар | 10 |
| Тестовое давление, бар | 14.3 |
| Горячая сторона | |
| Температура воды на входе/выходе, °C | 90/70 |
| Скорость протока, м³/ч | 10. 67 67 |
| Перепад давления, бар | 0.21 |
| Холодная сторона | |
| Температура воды на входе/выходе, °C | 20/35 |
| Скорость протока, м³/ч | 13.95 |
| Перепад давления, бар | 0.41 |
| Модель | 287 кВт |
|---|---|
| Мощность, кВт | 287 |
| Площадь теплопередачи, м² | 1.1 |
| Материал пластин | нержавеющая сталь AISI 316L |
| Количество пластин, шт | 29 |
| Толщина пластин, мм | 0.5 |
| Вес пустого теплообменника, кг | 64.9 |
| Вес полного теплообменника, кг | 67.87 |
| Объем контура горячей/холодной стороны, л | 2.96 |
| Рабочая температура, °C | от -20 до +109 |
| Рабочее давление, бар | 10 |
| Тестовое давление, бар | 14. 3 3 |
| Горячая сторона | |
| Температура воды на входе/выходе, °C | 90/70 |
| Скорость протока, м³/ч | 12.65 |
| Перепад давления, бар | 0.23 |
| Холодная сторона | |
| Температура воды на входе/выходе, °C | 20/35 |
| Скорость протока, м³/ч | 13.55 |
| Перепад давления, бар | 0.44 |
| Модель | 446 кВт |
|---|---|
| Мощность, кВт | 446 |
| Площадь теплопередачи, м² | 1.68 |
| Материал пластин | нержавеющая сталь AISI 316L |
| Количество пластин, шт | 29 |
| Толщина пластин, мм | 0.5 |
| Вес пустого теплообменника, кг | 68. 78 78 |
| Вес полного теплообменника, кг | 67.87 |
| Объем контура горячей/холодной стороны, л | 4.54 |
| Рабочая температура, °C | от -20 до +109 |
| Рабочее давление, бар | 10 |
| Тестовое давление, бар | 14.3 |
| Горячая сторона | |
| Температура воды на входе/выходе, °C | 90/70 |
| Скорость протока, м³/ч | 19.66 |
| Перепад давления, бар | 0.26 |
| Холодная сторона | |
| Температура воды на входе/выходе, °C | 20/35 |
| Скорость протока, м³/ч | 25.71 |
| Перепад давления, бар | 0.49 |
| Модель | 672 кВт |
|---|---|
| Мощность, кВт | 672 |
| Площадь теплопередачи, м² | 2. 37 37 |
| Материал пластин | нержавеющая сталь AISI 316L |
| Количество пластин, шт | 29 |
| Толщина пластин, мм | 0.5 |
| Вес пустого теплообменника, кг | 113.93 |
| Вес полного теплообменника, кг | 120.83 |
| Объем контура горячей/холодной стороны, л | 6.9 |
| Рабочая температура, °C | от -20 до +109 |
| Рабочее давление, бар | 10 |
| Тестовое давление, бар | 14.3 |
| Горячая сторона | |
| Температура воды на входе/выходе, °C | 90/70 |
| Скорость протока, м³/ч | 29.63 |
| Перепад давления, бар | 0.27 |
| Холодная сторона | |
| Температура воды на входе/выходе, °C | 20/35 |
| Скорость протока, м³/ч | 38. 74 74 |
| Перепад давления, бар | 0.48 |
| Модель | 819 кВт |
|---|---|
| Мощность, кВт | 819 |
| Площадь теплопередачи, м² | 2.99 |
| Материал пластин | нержавеющая сталь AISI 316L |
| Количество пластин, шт | 29 |
| Толщина пластин, мм | 0.5 |
| Вес пустого теплообменника, кг | 118.22 |
| Вес полного теплообменника, кг | 126.92 |
| Объем контура горячей/холодной стороны, л | 8.7 |
| Рабочая температура, °C | от -20 до +109 |
| Рабочее давление, бар | 10 |
| Тестовое давление, бар | 14.3 |
| Горячая сторона | |
| Температура воды на входе/выходе, °C | 90/70 |
| Скорость протока, м³/ч | 36. 11 11 |
| Перепад давления, бар | 0.28 |
| Холодная сторона | |
| Температура воды на входе/выходе, °C | 20/35 |
| Скорость протока, м³/ч | 47.22 |
| Перепад давления, бар | 0.49 |
Свернуть таблицу Развернуть таблицу
Ламеллярный ихтиоз: MedlinePlus Genetics
Описание
Ламеллярный ихтиоз — это заболевание, которое в основном поражает кожу. Младенцы с этим заболеванием обычно рождаются с плотной прозрачной оболочкой, покрывающей их кожу, называемой коллодиевой мембраной. Эта оболочка обычно высыхает и отслаивается в течение первых нескольких недель жизни, а затем становится очевидным, что пораженные дети имеют шелушащуюся кожу, а веки и губы вывернуты наружу. Люди с ламеллярным ихтиозом обычно имеют большие, темные, пластинчатые чешуйки, покрывающие кожу на большей части тела. У младенцев с ламеллярным ихтиозом могут развиться инфекции, чрезмерная потеря жидкости (обезвоживание) и проблемы с дыханием. У больных также может наблюдаться выпадение волос (алопеция), аномальное формирование ногтей на руках и ногах (дистрофия ногтей), снижение потоотделения (гипогидроз), повышенная чувствительность к теплу и утолщение кожи на ладонях и подошвах. стоп (кератодермия). Реже у больных наблюдается покраснение кожи (эритема) и деформация суставов (контрактуры).
У больных также может наблюдаться выпадение волос (алопеция), аномальное формирование ногтей на руках и ногах (дистрофия ногтей), снижение потоотделения (гипогидроз), повышенная чувствительность к теплу и утолщение кожи на ладонях и подошвах. стоп (кератодермия). Реже у больных наблюдается покраснение кожи (эритема) и деформация суставов (контрактуры).
Частота
Ламеллярный ихтиоз, по оценкам, поражает 1 из 100 000 человек в Соединенных Штатах. Это заболевание чаще встречается в Норвегии, где, по оценкам, им страдает 1 человек из 91 000.
Причины
Мутации в одном из многих генов могут вызывать ламеллярный ихтиоз. Эти гены обеспечивают инструкции по производству белков, которые находятся в самом внешнем слое кожи (эпидермисе). Аномалии кожи, связанные с ламеллярным ихтиозом, нарушают нормальное формирование эпидермиса, что приводит к нарушению регуляции температуры тела, задержке воды и устойчивости к инфекциям.
Мутации в гене TGM1 ответственны примерно за 90 процентов случаев ламеллярного ихтиоза. Ген TGM1 предоставляет инструкции по созданию фермента, называемого трансглютаминазой 1. Этот фермент участвует в формировании ороговевшей клеточной оболочки, которая представляет собой структуру, окружающую клетки кожи и помогающую сформировать защитный барьер между телом и окружающей средой. Мутации гена TGM1 приводят к резкому снижению или отсутствию продукции ферментов, что препятствует образованию ороговевшей клеточной оболочки.
Ген TGM1 предоставляет инструкции по созданию фермента, называемого трансглютаминазой 1. Этот фермент участвует в формировании ороговевшей клеточной оболочки, которая представляет собой структуру, окружающую клетки кожи и помогающую сформировать защитный барьер между телом и окружающей средой. Мутации гена TGM1 приводят к резкому снижению или отсутствию продукции ферментов, что препятствует образованию ороговевшей клеточной оболочки.
Мутации в других генах, связанных с ламеллярным ихтиозом, ответственны лишь за небольшой процент случаев. У некоторых людей с ламеллярным ихтиозом причина заболевания неизвестна. Исследователи идентифицировали несколько участков хромосом, которые содержат гены, которые могут быть связаны с ламеллярным ихтиозом, хотя конкретные гены не были идентифицированы.
Наследование
Это состояние наследуется по аутосомно-рецессивному типу, что означает, что обе копии гена в каждой клетке имеют мутации. Каждый из родителей человека с аутосомно-рецессивным заболеванием несет по одной копии мутировавшего гена, но обычно у них нет признаков и симптомов заболевания.
Другие названия для этого состояния
- Коллодиевый детский
- Коллодиевый детский синдром
- Ихтиоз пластинчатый
- Ихтиоз пластинчатый
- LI
Дополнительная информация и ресурсы
Информация о генетическом тестировании
- Реестр генетического тестирования: аутосомно-рецессивный врожденный ихтиоз 3
- Реестр генетического тестирования: аутосомно-рецессивный врожденный ихтиоз 4A
- Реестр генетического тестирования: аутосомно-рецессивный врожденный ихтиоз 5
- Реестр генетического тестирования: аутосомно-рецессивный врожденный ихтиоз 8
- Реестр генетического тестирования: врожденный ихтиоз кожи
Информационный центр генетических и редких заболеваний
- Ламеллярный ихтиоз
Ресурсы поддержки пациентов и защиты интересов
- Информационный поиск по болезни
- Национальная организация редких заболеваний (NORD)
Научные исследования от ClinicalTrials.
 gov
gov- ClinicalTrials.gov
Каталог генов и болезней от OMIM
- ИХТИОЗ ВРОЖДЕННЫЙ АУТОСОМНО-РЕЦЕССНЫЙ 1
- ИХТИОЗ ВРОЖДЕННЫЙ АУТОСОМНО-РЕЦЕССОВЫЙ 3
- ИХТИОЗ ВРОЖДЕННЫЙ АУТОСОМНО-РЕЦЕССОВЫЙ 4A
- ИХТИОЗ ВРОЖДЕННЫЙ АУТОСОМНО-РЕЦЕССОВЫЙ 5
Научные статьи в PubMed
- ПабМед
Ссылки
- Akiyama M. Мутации ABCA12 и аутосомно-рецессивный врожденный ихтиоз: a обзор корреляций генотип/фенотип и патогенетических концепций. Гул Мутат. 2010 г.; 31 октября (10): 1090-6. doi: 10.1002/humu.21326. Обзор. Цитата в PubMed
- Фарасат С., Вэй М.Х., Герман М., Ливер Д.Дж., Стейнберг С.М., Бэйл С.Дж., Флекман П.,
Торо младший. Новые мутации трансглютаминазы-1 и исследования генотип-фенотип
из 104 пациентов с аутосомно-рецессивным врожденным ихтиозом в США. Джей Мед
Жене. 2009 г.Фев; 46 (2): 103-11. doi: 10.
 1136/jmg.2008.060905. Epub 2008 Oct 23. Цитирование в PubMed или бесплатная статья на PubMed Central
1136/jmg.2008.060905. Epub 2008 Oct 23. Цитирование в PubMed или бесплатная статья на PubMed Central - Герман М.Л., Фарасат С., Штайнбах П.Дж., Вэй М.Х., Туре О., Флекман П., Блейк П., Бэйл SJ, Торо JR. Мутации гена трансглютаминазы-1 при аутосомно-рецессивном врожденном ихтиоз: сводка мутаций (в том числе 23 новых) и моделирование TGase-1. Хум Мутат. 2009 апр; 30 (4): 537-47. doi: 10.1002/humu.20952. Обзор. Цитирование в PubMed или бесплатная статья в PubMed Central
- Исраэл С., Хамайси З., Фукс-Телем Д., Нусбек Дж., Бергман Р., Сариг О., Спрехер E. Мутация в LIPN, кодирующем эпидермальную липазу N, вызывает форму позднего начала аутосомно-рецессивный врожденный ихтиоз. Am J Hum Genet. 2011 апр 8;88(4):482-7. doi: 10.1016/j.ajhg.2011.02.011. Цитирование в PubMed или бесплатная статья в PubMed Central
- Одзи В., Тадини Г., Акияма М., Бланше Бардон С., Бодемер С., Бурра Э., Кудьер
П., ДиДжиованна Дж.Дж., Элиас П.
 , Фишер Дж., Флекман П., Джина М., Харпер Дж., Хашимото Т.,
Хауссер И., Хеннис Х.К., Холь Д., Овнанян А., Исида-Ямамото А., Яцик В.К., Личман
С., Ли И., Мазереу-Отье Дж., Милстон Л., Морис-Пикард Ф., Паллер А.С., Ричард
Г., Шмут М., Симидзу Х., Спрехер Э., Ван Стенсел М., Тайеб А., Торо Дж. Р., Вабрес П.,
Валквист А., Уильямс М., Трауп Х. Пересмотренная номенклатура и классификация
наследственные ихтиозы: результаты Первой консенсусной конференции по ихтиозам в
Сорез 2009. J Am Acad Дерматол. 2010 Октябрь; 63 (4): 607-41. дои:
10.1016/j.jaad.2009.11.020. Обзор. Цитата в PubMed
, Фишер Дж., Флекман П., Джина М., Харпер Дж., Хашимото Т.,
Хауссер И., Хеннис Х.К., Холь Д., Овнанян А., Исида-Ямамото А., Яцик В.К., Личман
С., Ли И., Мазереу-Отье Дж., Милстон Л., Морис-Пикард Ф., Паллер А.С., Ричард
Г., Шмут М., Симидзу Х., Спрехер Э., Ван Стенсел М., Тайеб А., Торо Дж. Р., Вабрес П.,
Валквист А., Уильямс М., Трауп Х. Пересмотренная номенклатура и классификация
наследственные ихтиозы: результаты Первой консенсусной конференции по ихтиозам в
Сорез 2009. J Am Acad Дерматол. 2010 Октябрь; 63 (4): 607-41. дои:
10.1016/j.jaad.2009.11.020. Обзор. Цитата в PubMed - Родригес-Пасос Л., Хинарте М., Вега А., Торибио Дж. Аутосомно-рецессивный врожденный ихтиоз. Actas Дермосифилиогр. 2013 май; 104(4):270-84. дои: 10.1016/j.adengl.2011.11.021. Epub 2013 3 апр. Обзор. Английский испанский. Цитата в PubMed
- Терринони А., Серра В., Кодиспоти А., Таламонти Э., Буй Л., Паломбо Р., Сетте М.,
Кампионе Э., Дидона Б., Анниккиарико-Петруцелли М.
 , Самбруно Г., Мелино Г., Канди Э.
Новые мутации трансглютаминазы 1 у пациентов с ламеллярным ихтиозом.
Клеточная смерть Дис. 2012, 25 октября; 3:e416. doi: 10.1038/cddis.2012.152. Цитирование в PubMed или бесплатная статья в PubMed Central
, Самбруно Г., Мелино Г., Канди Э.
Новые мутации трансглютаминазы 1 у пациентов с ламеллярным ихтиозом.
Клеточная смерть Дис. 2012, 25 октября; 3:e416. doi: 10.1038/cddis.2012.152. Цитирование в PubMed или бесплатная статья в PubMed Central
Ламеллярный ихтиоз Информация | Гора Синай
ЛИ; Коллодий бэби — ламеллярный ихтиоз; врожденный ихтиоз; Аутосомно-рецессивный врожденный ихтиоз — ламеллярный ихтиоз тип
Ламеллярный ихтиоз (ЛИ) — редкое кожное заболевание. Он появляется при рождении и сохраняется на протяжении всей жизни.
Ихтиоз возникает в результате аномалии ороговения (образования наружного омертвевшего слоя) кожи. Различают множество форм ихтиоза. Обратите внимание на сухую кожу и шелушение. Ихтиоз обычно ухудшается зимой и сильнее поражает ноги.
Различают множество форм ихтиоза. Обратите внимание на сухую кожу и шелушение. Ихтиоз обычно ухудшается зимой и сильнее поражает ноги.
Причины
ЛИ — это аутосомно-рецессивное заболевание. Это означает, что и мать, и отец должны передать своему ребенку одну аномальную копию гена болезни, чтобы у ребенка развилась болезнь.
Симптомы
Многие дети с ЛИ рождаются с прозрачным, блестящим, восковым слоем кожи, называемым коллодиевой мембраной. По этой причине эти младенцы известны как коллодиевые младенцы. Мембрана сбрасывается в течение первых 2 недель жизни. Кожа под мембраной красная и чешуйчатая, напоминающая поверхность рыбы.
При ЛН внешний слой кожи, называемый эпидермисом, не может защитить тело так, как это может сделать здоровый эпидермис. В результате у ребенка с ЛИ могут возникнуть следующие проблемы со здоровьем:
В результате у ребенка с ЛИ могут возникнуть следующие проблемы со здоровьем:
- Затруднения при кормлении
- Потеря жидкости (обезвоживание)
- Потеря баланса минералов в организме (электролитный дисбаланс)
- Проблемы с дыханием
- Организм нестабильная температура
- Инфекции кожи или всего тела
У детей старшего возраста и взрослых с ЛИ могут быть следующие симптомы:
- Гигантские чешуйки, покрывающие большую часть тела
- Снижение потоотделения, вызывающее чувствительность к теплу
- Выпадение волос
- Аномальные ногти пальцев рук и ног
- Кожа ладоней и подошв утолщена
Лечение
Детям с коллодием обычно необходимо оставаться в отделении интенсивной терапии новорожденных (ОИТН). Их помещают в инкубатор с повышенной влажностью. Им потребуются дополнительные подкормки. На кожу нужно наносить увлажняющие средства. После отслоения коллодиевой мембраны младенцев обычно можно отправлять домой.
Их помещают в инкубатор с повышенной влажностью. Им потребуются дополнительные подкормки. На кожу нужно наносить увлажняющие средства. После отслоения коллодиевой мембраны младенцев обычно можно отправлять домой.
Уход за кожей на протяжении всей жизни заключается в том, чтобы поддерживать кожу во влажном состоянии, чтобы свести к минимуму толщину чешуек. Меры включают:
- Увлажняющие средства, наносимые на кожу
- Лекарства, называемые ретиноидами, которые принимаются перорально в тяжелых случаях
- Среда с высокой влажностью
- Купание для удаления чешуи
Возможные осложнения
Младенцы подвергаются риску заражения, когда они теряют коллодиевую мембрану.
Проблемы с глазами могут возникнуть в более позднем возрасте, поскольку глаза не могут полностью закрыться.
Мартин К.Л. Нарушения кератинизации. В: Клигман Р.М., Сент-Геме Дж.В., Блюм Н.Дж., Шах С.С. Таскер Р.К., Уилсон К.М., ред. Учебник педиатрии Нельсона . 21-е изд. Филадельфия, Пенсильвания: Elsevier; 2020: глава 677.
Паттерсон Дж.В. Нарушения созревания и ороговения эпидермиса. В: Паттерсон Дж. В., изд. Кожная патология Уидона . 5-е изд. Филадельфия, Пенсильвания: Elsevier; 2021: глава 10.
Ричард Г., Рингпфейл Ф. Ихтиозы, эритрокератодермия и родственные заболевания. В: Болонья Дж. Л., Шаффер Дж. В., Черрони Л., ред. Дерматология . 4-е изд. Филадельфия, Пенсильвания: Elsevier; 2018: глава 57.
Последнее рассмотрение: 10.11.2020
Рецензию сделал: Рамин Фатхи, доктор медицинских наук, FAAD, директор Phoenix Surgical Dermatology Group, Феникс, Аризона. Также рассмотрены Дэвидом Зивом, доктором медицины, MHA, медицинским директором, Брендой Конауэй, редакционным директором, и A.D.A.M. Редакционная коллегия.
Также рассмотрены Дэвидом Зивом, доктором медицины, MHA, медицинским директором, Брендой Конауэй, редакционным директором, и A.D.A.M. Редакционная коллегия.
Пластинчатый ихтиоз — NORD (Национальная организация по редким заболеваниям)
Ламеллярный ихтиоз
NORD выражает благодарность Пери Л. Чавусгил, MSc, стажеру редактора NORD из Программы обучения генетическому консультированию Университета Эмори, и Сесилии А. Беллкросс, доктору философии, магистру наук, CGC , доценту, директору учебной программы генетического консультирования Медицинской школы Университета Эмори за помощь в подготовке этого отчета.
Синонимы ламеллярного ихтиоза
- коллодий детский
- врожденный ламеллярный ихтиоз
- LI
Признаки и симптомы
Причины
LI может быть вызван вредными изменениями в нескольких генах. Наиболее распространенным геном, связанным с этим заболеванием, является TGM1 . Другие гены включают NIPAL4 , ALOX12B и CYP4F22 . Эти гены дают инструкции по производству ферментов и белков. Эти ферменты и белки важны для нормального развития, функционирования и отторжения клеток кожи. Имеются также данные о том, что с LI связано больше генов. Тяжесть состояния не всегда может быть определена на основании результатов генетического тестирования.
Эти гены дают инструкции по производству ферментов и белков. Эти ферменты и белки важны для нормального развития, функционирования и отторжения клеток кожи. Имеются также данные о том, что с LI связано больше генов. Тяжесть состояния не всегда может быть определена на основании результатов генетического тестирования.
Вредные изменения, вызывающие ЛН, наследуются рецессивно. Рецессивные генетические нарушения возникают, когда человек наследует нерабочий ген от каждого родителя. Если человек получает один рабочий ген и один неработающий ген болезни, он будет носителем болезни, но обычно не будет проявлять симптомов. Риск для двух родителей-носителей передать неработающий ген и, следовательно, родить больного ребенка составляет 25% при каждой беременности. Риск рождения ребенка-носителя, как и у родителей, составляет 50% при каждой беременности. Вероятность того, что ребенок получит рабочие гены от обоих родителей, составляет 25%. Риск одинаков для мужчин и женщин.
Затронутые группы населения
LI встречается примерно у одного из 200 000 человек. Заболевание может затронуть людей любого пола, расы и этнической принадлежности.
Заболевание может затронуть людей любого пола, расы и этнической принадлежности.
Стандартная терапия
Лечение
LI обычно лечат местно. Врачи используют кремы, которые помогают восстановить кожный барьер. Эти кремы часто содержат керамиды или холестерин. Также можно использовать увлажняющие средства с вазелином или ланолином. Иногда в качестве вариантов лечения используются легкие кератолитики или ретиноиды для местного применения. Врачи могут лечить тяжелую НЛ пероральными ретиноидами. Ретиноиды могут быть токсичными для организма, поэтому их применяют с осторожностью.
Для диагностики и лечения ARCI, включая ЛИ, людям может потребоваться консультация следующих врачей-специалистов:
- Клинический генетик и/или генетик-консультант
- Кожный врач (дерматолог), знакомый с врожденным ихтиозом
- Окулист (офтальмолог), знакомый при врожденном ихтиозе
Могут потребоваться анализы на проблемы с терморегуляцией и потерю воды, а также тесты на инфекции.
Исследовательская терапия
Большинство современных методов лечения не устраняют воспаление кожи, которое наблюдается у некоторых людей с ЛИ. Исследователи изучают использование противовоспалительных препаратов, используемых для лечения псориаза (другой тип кожного заболевания) у людей с ЛИ. Исследователи также пытаются персонализировать лечение для конкретного вредного изменения гена, которое есть у пациента. Этот подход может привести к улучшению лечения людей с LI. Наконец, проводятся экспериментальные исследования, чтобы определить, может ли ферментная заместительная терапия (ФЗТ) быть вариантом лечения. Однако ФЗТ будет вариантом только для людей с определенными вредными генными изменениями.
Информация о текущих клинических испытаниях размещена в Интернете по адресу https://clinicaltrials.gov/. Все исследования, финансируемые правительством США, а некоторые из них поддерживаются частным сектором, публикуются на этом правительственном веб-сайте.
Для получения информации о клинических испытаниях, проводимых в Клиническом центре NIH в Бетесде, штат Мэриленд, обращайтесь в отдел набора пациентов NIH:
Бесплатный звонок: (800) 411-1222
Телетайп: (866) 411-1010
Электронная почта: [email protected]
Некоторые текущие клинические испытания также размещены на следующей странице веб-сайта NORD:
https://rarediseases. org/for-patients-and-families/information-resources/info-clinical-trials-and-research-studies/
org/for-patients-and-families/information-resources/info-clinical-trials-and-research-studies/
Для получения информации о клинических испытаниях, спонсируемых частными источниками, обращайтесь:
http: //www.centerwatch.com/
Для получения информации о клинических испытаниях, проведенных в Европе, обращайтесь по телефону:
https://www.clinicaltrialsregister.eu/
Для получения информации о текущих исследованиях, связанных с ихтиозом, обращайтесь в Фонд ихтиоза и родственных заболеваний Типы скинов, перечисленные в разделе «Ресурсы» данного отчета.
Ссылки
СТАТЬИ В ЖУРНАЛЕ
Валквист А., Фишер Дж., Торме Х. Унаследованные несиндромальные ихтиозы: обновленная информация о патофизиологии, диагностике и лечении. Am J Clin Дерматол. 2018;19(1):51-66.
Марукян Н.В., Чоатэ К.А. Последние достижения в понимании патогенеза ихтиоза. F1000рез. 2016;5.
Такеичи Т., Акияма М. Наследственный ихтиоз: несиндромальные формы. Журнал дерматологии. 2016;43(3):242-251.
2016;43(3):242-251.
Дайер Дж.А., Спракер М., Уильямс М. Уход за новорожденным с ихтиозом. Дерматологическая терапия. 2013;26(1):1-15.
Chaurasia S, Das S, Ramamurthy B. Микробный кератит при пластинчатом ихтиозе. Инт офтальмол. 2008;28(5):367-368.
Смолл К., Гинзбург Х., Греко М.А., Сарита-Рейес С., Купчик Г., Блей Ф. Больше, чем глубоко на коже: случай врожденного ламеллярного ихтиоза, лимфатической мальформации и других аномалий. Лимфатические исследования и биология. 2008;6(1):39-44.
Кумар Т.С., Скотт XJ, Саймон А., Рагхупати П. Дефицит витамина D рахит с ламеллярным ихтиозом. Журнал последипломной медицины. 2007;53(3):215.
DiGiovanna JJ, Robinson-Bostom L. Ихтиоз: этиология, диагностика и лечение. Американский журнал клинической дерматологии. 2003;4(2):81.
Уильямс М.Л., Элиас П.М. Просвещенная терапия нарушений ороговения. Клиники дерматологии. 2003;21(4):269-273.
Уильямс МЛ. Ламеллярный ихтиоз и врожденная ихтиозиформная эритродермия (CIE). Ихтиоз Фокус. Осень 2003 г.; 22(3): 1-4.
Ихтиоз Фокус. Осень 2003 г.; 22(3): 1-4.
Баксман М., Хикман Дж., Рэгсдейл В., Носилки Г., Крохмаль Л., Вер РФ. Терапевтическая активность лактатного 12% лосьона при лечении ихтиоза: активен по сравнению с носителем и активен по сравнению с вазелиновым кремом. Журнал Американской академии дерматологии. 1986;15(6):1253-1258.
ИНТЕРНЕТ
Ричард Г. Аутосомно-рецессивный врожденный ихтиоз. 10 января 2001 г. [Обновлено 18 мая 2017 г.]. В: Адам М.П., Ардингер Х.Х., Пагон Р.А. и др., редакторы. GeneReviews® [Интернет]. Сиэтл (Вашингтон): Вашингтонский университет, Сиэтл; 1993-2020 гг. Доступно по адресу: https://www.ncbi.nlm.nih.gov/books/NBK1420/ По состоянию на 6 июля 2020 г.
Годы публикации
2005, 2020
Информация в базе данных NORD по редким заболеваниям предназначена только для образовательных целей. и не предназначен для замены консультации врача или другого квалифицированного медицинского работника.
Содержание веб-сайта и баз данных Национальной организации редких заболеваний (NORD) защищено авторским правом и не может быть воспроизведено, скопировано, загружено или распространено каким-либо образом в коммерческих или общественных целях без предварительного письменного разрешения и одобрения.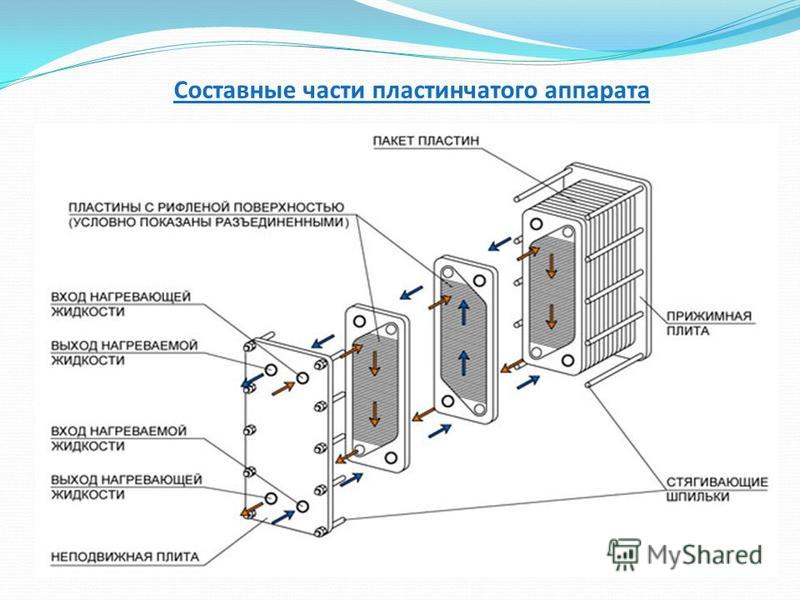 от НОРД. Физические лица могут распечатать одну бумажную копию отдельного заболевания для личного использования при условии, что содержание не изменено и включает авторские права NORD.
от НОРД. Физические лица могут распечатать одну бумажную копию отдельного заболевания для личного использования при условии, что содержание не изменено и включает авторские права NORD.
Национальная организация редких заболеваний (NORD)
55 Kenosia Ave., Danbury CT 06810 • (203)744-0100
Посткорреляционная крио-CLEM на ламеллах выявляет мембранную архитектуру ламеллярных тел альвеолярные клетки. LB разбираются в липидно-белковую сеть, которая снижает поверхностное натяжение и облегчает газообмен в альвеолярной полости. Текущие знания об архитектуре LB преимущественно основаны на исследованиях электронной микроскопии с использованием разрушающих методов подготовки образцов. Мы установили и утвердили подход посткорреляционной криокорреляционной световой и электронной микроскопии на ламеллах для измельченных крио-FIB клеток для структурной характеристики и подтверждения идентичности LB в их невозмущенном состоянии. Используя деконволюцию и регистрацию 3D-изображений, мы смогли идентифицировать флуоресцентно меченные мембранные структуры, проанализированные с помощью криоэлектронной томографии.
 Криоэлектронная томография in situ A549клетки, а также первичные эпителиальные клетки малых дыхательных путей человека показали, что LB состоят из мембранных листов, часто прикрепленных к пограничной мембране через Т-образные соединения. Мы сообщаем о до сих пор неописанном белковом комплексе купола наружной мембраны (OMDP) на ограничивающей мембране LBs. Наши данные предполагают, что биогенез LB управляется параллельным переносом мембранных листов и кривизной ограничивающей мембраны, чтобы максимизировать емкость хранения липидов.
Криоэлектронная томография in situ A549клетки, а также первичные эпителиальные клетки малых дыхательных путей человека показали, что LB состоят из мембранных листов, часто прикрепленных к пограничной мембране через Т-образные соединения. Мы сообщаем о до сих пор неописанном белковом комплексе купола наружной мембраны (OMDP) на ограничивающей мембране LBs. Наши данные предполагают, что биогенез LB управляется параллельным переносом мембранных листов и кривизной ограничивающей мембраны, чтобы максимизировать емкость хранения липидов.Введение
Пластинчатые тельца (LB) представляют собой специализированные органеллы, обнаруженные исключительно в эпителиальных клетках альвеолярного типа 2 (AEC2) и в кератиноцитах 1 . Альвеолярные LB продуцируют, накапливают и секретируют сурфактант, смесь специализированных липидов и белков. При секреции в альвеолярную полость он быстро распадается на высокоорганизованную сеть. Легочный сурфактант снижает поверхностное натяжение на границе воздух-вода в альвеолах, облегчая газообмен во время дыхания. Поэтому его необходимо постоянно пополнять для поддержания дыхания 2 . AEC2 и сурфактант являются основными участниками легочного иммунного ответа 3 . Дефекты выработки сурфактанта связаны с повышенным риском респираторных инфекций, вызванных такими патогенами, как вирус гриппа А 4 , респираторно-синцитиальный вирус 5 , пневмония 6 и Mycobacterium tuberculosis 7 . Сурфактантный белок D (SP-D) специфически связывает гликозилированные патогены, в том числе SARS-CoV-1 8 . Метаболизм жирных кислот AEC2 и ультраструктура LB серьезно нарушены пандемическим штаммом гриппа h2N1 9 и высокопатогенный штамм H7N9 10 . Несмотря на их важность для здоровья и болезней, многие вопросы о биогенезе, структуре и секреции LB остаются открытыми. LB состоят из ядра, содержащего многослойные мембранные листы, окруженные ограничивающей мембраной, как показано с помощью просвечивающей электронной микроскопии тонких срезов (ПЭМ) 11 .
Поэтому его необходимо постоянно пополнять для поддержания дыхания 2 . AEC2 и сурфактант являются основными участниками легочного иммунного ответа 3 . Дефекты выработки сурфактанта связаны с повышенным риском респираторных инфекций, вызванных такими патогенами, как вирус гриппа А 4 , респираторно-синцитиальный вирус 5 , пневмония 6 и Mycobacterium tuberculosis 7 . Сурфактантный белок D (SP-D) специфически связывает гликозилированные патогены, в том числе SARS-CoV-1 8 . Метаболизм жирных кислот AEC2 и ультраструктура LB серьезно нарушены пандемическим штаммом гриппа h2N1 9 и высокопатогенный штамм H7N9 10 . Несмотря на их важность для здоровья и болезней, многие вопросы о биогенезе, структуре и секреции LB остаются открытыми. LB состоят из ядра, содержащего многослойные мембранные листы, окруженные ограничивающей мембраной, как показано с помощью просвечивающей электронной микроскопии тонких срезов (ПЭМ) 11 . ЛБ содержат 85% фосфолипидов по массе, в основном дипальмитоилфосфатидилхолин (ДПФХ), ненасыщенные фосфатидилхолины, фосфатидилглицерины 12 , а также холестерин и специализированные сурфактантные белки A, B и C (SP-A, SP-B и SP-C) 13 . Большинство белков, связанных с LB, обычно находятся в лизосомах, поэтому LB классифицируются как органеллы, связанные с лизосомами. Масс-спектрометрия идентифицировала 34 белка, уникальных для LB легких 13 . В то время как сердцевина содержит малые гидрофобные белки SP-B и SP-C 14 , ограничивающая мембрана обогащена флиппазой АТФ-связывающей кассеты подсемейства A 3 (ABCA3) 15 .
ЛБ содержат 85% фосфолипидов по массе, в основном дипальмитоилфосфатидилхолин (ДПФХ), ненасыщенные фосфатидилхолины, фосфатидилглицерины 12 , а также холестерин и специализированные сурфактантные белки A, B и C (SP-A, SP-B и SP-C) 13 . Большинство белков, связанных с LB, обычно находятся в лизосомах, поэтому LB классифицируются как органеллы, связанные с лизосомами. Масс-спектрометрия идентифицировала 34 белка, уникальных для LB легких 13 . В то время как сердцевина содержит малые гидрофобные белки SP-B и SP-C 14 , ограничивающая мембрана обогащена флиппазой АТФ-связывающей кассеты подсемейства A 3 (ABCA3) 15 .
В современной модели биогенеза LB липиды переворачиваются ABCA3 из цитозольного листка в просвет и импортируются в ядро LB 16 , где SP-B и SP-C ответственны за дальнейшую перестройку липидов в плотно упакованные мембранные листы 17 . Однако эту модель было трудно проверить. Высокое содержание липидов в LB плохо сохраняется в результате подготовки образцов TEM при комнатной температуре, которая основана на химической фиксации и обезвоживании. В результате концентрические мембраны внутри LB кажутся морщинистыми. Следовательно, неизвестно, как они организованы в трех измерениях (3D), и неизвестно, как формируются стопки мембран. Исследование с использованием криоэлектронной микроскопии (ЭМ) витрифицированных срезов (CEMOVIS) на легких крыс позволило визуализировать замороженные гидратированные LB и показало гладкие концентрические мембраны 18 . Однако из-за артефактов сжатия, вызванных секциями, и отсутствия совместимости с криоэлектронной томографией (крио-ЭТ) 19 исследование дало лишь небольшое представление о сложной архитектуре LB.
В результате концентрические мембраны внутри LB кажутся морщинистыми. Следовательно, неизвестно, как они организованы в трех измерениях (3D), и неизвестно, как формируются стопки мембран. Исследование с использованием криоэлектронной микроскопии (ЭМ) витрифицированных срезов (CEMOVIS) на легких крыс позволило визуализировать замороженные гидратированные LB и показало гладкие концентрические мембраны 18 . Однако из-за артефактов сжатия, вызванных секциями, и отсутствия совместимости с криоэлектронной томографией (крио-ЭТ) 19 исследование дало лишь небольшое представление о сложной архитектуре LB.
В отличие от CEMOVIS, измельчение с помощью криофокусированного ионного пучка (крио-ФИБ) позволяет препарировать тонкие ячеистые ламели произвольной толщины с гладкой поверхностью и без сжатия, так что они совместимы с крио-ЕТ 20,21 . Корреляционный свет и ЭМ (CLEM) позволяют однозначно идентифицировать целевые отсеки и дают структурные детали 22 . Методы CLEM были адаптированы для крио-ЭМ и успешно применялись на образцах in vitro 23,24 или для крио-ЭТ на целых клетках 25,26,27,28 . Однако корреляция данных световой микроскопии (LM) и ЭМ в рабочем процессе, включающем измельчение крио-FIB, затруднена из-за геометрии измельчения и многократного переноса между микроскопами: каждый перенос увеличивает риск расстеклования образца и загрязнения льдом. На данный момент доступные рабочие процессы крио-CLEM in situ, включающие измельчение крио-ФИБ, нацелены на измельчение крио-ФИБ в конкретных местах 29,30 , но не предлагают целевую проверку.
Методы CLEM были адаптированы для крио-ЭМ и успешно применялись на образцах in vitro 23,24 или для крио-ЭТ на целых клетках 25,26,27,28 . Однако корреляция данных световой микроскопии (LM) и ЭМ в рабочем процессе, включающем измельчение крио-FIB, затруднена из-за геометрии измельчения и многократного переноса между микроскопами: каждый перенос увеличивает риск расстеклования образца и загрязнения льдом. На данный момент доступные рабочие процессы крио-CLEM in situ, включающие измельчение крио-ФИБ, нацелены на измельчение крио-ФИБ в конкретных местах 29,30 , но не предлагают целевую проверку.
Здесь мы показываем, что точное знание положения ламеллы в контексте всей клетки, определенное с помощью крио-ЛМ после крио-ТЭМ, облегчает точное сопоставление исходных данных ЛМ с клеточными структурами на ламелле. В представленном рабочем процессе двумерная корреляция применяется для нацеливания интересующей области на фрезерование в плоскости X–Y сетки. Второй этап корреляции с использованием данных LM, полученных после крио-ПЭМ-изображения, деконволюции и 3D-корреляции, затем применяется для идентификации наблюдаемых структур, соответствующих положению ламели не только в X – Y, но и в Z-измерении. Мы показываем, что последнее необходимо для повышения точности корреляции путем компьютерного удаления флуоресцентных сигналов вне ламеллы.
Второй этап корреляции с использованием данных LM, полученных после крио-ПЭМ-изображения, деконволюции и 3D-корреляции, затем применяется для идентификации наблюдаемых структур, соответствующих положению ламели не только в X – Y, но и в Z-измерении. Мы показываем, что последнее необходимо для повышения точности корреляции путем компьютерного удаления флуоресцентных сигналов вне ламеллы.
Мы применили посткорреляционный рабочий процесс крио-CLEM на пластинке для изучения LB в клетках A549, модели для AEC2 31 , которые были временно трансфицированы ABCA3-eGFP, хорошо охарактеризованным маркером LB 15 . После обоих этапов корреляции 76% сигнала ABCA3-eGFP соответствовали мембраносвязанным органеллам, содержащим либо везикулы, либо ламеллярные мембраны, типичные для LB. Крио-ЭТ in situ позволила нам структурно охарактеризовать организацию мембран в ABCA3-eGFP-позитивных LB без артефактов подготовки образцов. Сердцевина LB представляет собой плотно упакованные листы мембраны с различной кривизной. Мы обнаружили параллельные двухслойные листы, соединенные перпендикулярно пограничной мембране через Т-образные соединения, и концентрические двухслойные листы как характерные структуры LB. Кроме того, наша работа выявила большой купольный белок наружной мембраны (OMDP) на пограничной мембране некоторых LB, предположительно участвующий в их формировании и транспортировке. Чтобы подтвердить наши выводы, мы проанализировали LB в первичных клетках легких человека, где мы наблюдали как «T»-переходы, так и OMDP.
Мы обнаружили параллельные двухслойные листы, соединенные перпендикулярно пограничной мембране через Т-образные соединения, и концентрические двухслойные листы как характерные структуры LB. Кроме того, наша работа выявила большой купольный белок наружной мембраны (OMDP) на пограничной мембране некоторых LB, предположительно участвующий в их формировании и транспортировке. Чтобы подтвердить наши выводы, мы проанализировали LB в первичных клетках легких человека, где мы наблюдали как «T»-переходы, так и OMDP.
Результаты
Разработка рабочего процесса пост-корреляции крио-CLEM на ламелях
Целью этого рабочего процесса корреляции является (i) его применимость для адгезивных клеток, выращенных непосредственно на крио-ЭМ сетках, (ii) быть совместим с коммерчески доступными установками крио-LM с широким полем зрения, и (iii) для обеспечения проверки цели путем корреляции флуоресцентного сигнала, полученного из положения ламели, с картой крио-ПЭМ ламеллы. Мотивация состоит в том, чтобы создать надежный метод крио-CLEM, который можно легко адаптировать. Для этого предоставляются все необходимые скрипты и программные плагины (см. раздел «Доступность кода»).
Для этого предоставляются все необходимые скрипты и программные плагины (см. раздел «Доступность кода»).
Цель этого рабочего процесса крио-CLEM на пластинке состоит в том, чтобы точно локализовать сигнал флуоресценции, полученный от пластинки, с картой крио-ТЕМ как в плоскости X–Y, так и в Z-измерении. В то время как первое является простым, второе является сложным, потому что (i) Z-размер ламеллы уменьшен до менее чем 5% от исходного Z-размера клетки (криоламеллы имеют толщину 150–200 нм, в то время как интактная клетка имеет толщину 4–6 мкм) и (ii) крио-LM имеет ограниченное Z-разрешение. Чтобы преодолеть эти трудности, мы внедрили рабочий процесс крио-CLEM с посткорреляцией на ламелле, в котором используются трехмерные карты крио-LM после деконволюции до и после подготовки крио-ламелей (рис. 1). Мы решили выполнить крио-LM получение крио-ламелей после крио-ТЭМ, так как изображение в крио-ЛМ увеличивает загрязнение льдом и, следовательно, снижает качество крио-ЭТ. Это приводит к потере флуоресценции на ламелях из-за повреждения электронного луча, происходящего во время крио-ЭТ. Поэтому мы использовали неотображаемое окружающее тело клетки, которое сохраняет свою флуоресценцию, для регистрации карты крио-ЛМ, полученной до и после получения крио-ТЭМ (дополнительный рисунок 1). Мы используем карту крио-ЛМ, полученную после крио-ТЭМ, для определения положения и наклона ламеллы, а исходную карту крио-ЛМ, полученную до измельчения, мы используем для получения сигнала флуоресценции. После 3D-регистрации двух крио-LM карт после деконволюции можно измерить наклон ламели с помощью сигнала светлого поля проходящего света (TL-BF) в плоскости Z – Y и соответствующим образом наклонить весь композитный пакет. Это позволяет нам извлечь один срез стека зарегистрированных и скорректированных по наклону изображений, содержащий только флуоресцентный сигнал, соответствующий пластинке. Используя жесткое преобразование, эту карту флуоресценции можно, наконец, сопоставить с картой крио-ТЕМ, что облегчает идентификацию интересующей структуры, анализируемой крио-ЭТ. Поскольку корреляция карт крио-ЛМ и крио-ТЭМ выполняется после крио-ЭТ, мы назвали этот подход «пост-корреляцией крио-CLEM на пластинке».
Поэтому мы использовали неотображаемое окружающее тело клетки, которое сохраняет свою флуоресценцию, для регистрации карты крио-ЛМ, полученной до и после получения крио-ТЭМ (дополнительный рисунок 1). Мы используем карту крио-ЛМ, полученную после крио-ТЭМ, для определения положения и наклона ламеллы, а исходную карту крио-ЛМ, полученную до измельчения, мы используем для получения сигнала флуоресценции. После 3D-регистрации двух крио-LM карт после деконволюции можно измерить наклон ламели с помощью сигнала светлого поля проходящего света (TL-BF) в плоскости Z – Y и соответствующим образом наклонить весь композитный пакет. Это позволяет нам извлечь один срез стека зарегистрированных и скорректированных по наклону изображений, содержащий только флуоресцентный сигнал, соответствующий пластинке. Используя жесткое преобразование, эту карту флуоресценции можно, наконец, сопоставить с картой крио-ТЕМ, что облегчает идентификацию интересующей структуры, анализируемой крио-ЭТ. Поскольку корреляция карт крио-ЛМ и крио-ТЭМ выполняется после крио-ЭТ, мы назвали этот подход «пост-корреляцией крио-CLEM на пластинке».
Рабочий процесс состоит из двух частей: сбор данных ( a – d ) и обработка данных для корреляционного анализа ( e – g ). Структуры в клетках, выращенных на ЭМ-сетках, флуоресцентно метят перед витрификацией методом глубокой заморозки. a На первом этапе витрифицированный образец картируется крио-LM, получая Z-стеки с интервалом 300 нм. б На втором этапе меченые клетки истончают путем измельчения крио-FIB для получения ламелей толщиной 150–200 нм. c Пластинки нанесены на карту с помощью крио-ПЭМ, и получены серии наклонных областей, представляющих интерес. d Области, отфрезерованные FIB, снова картируются с помощью крио-LM для определения точного положения ламелей. Для обоих Z-стеков шага ( a ) и шага ( d ) выполняется деконволюция для увеличения разрешения (не показано на рисунке). e Чтобы объединить информацию о флуоресцентном сигнале из шага ( a ) и положение ламели от шага ( d ), оба Z-стека выравниваются с помощью автоматизированного алгоритма 3D-регистрации. После регистрации изображения канал светлого поля проходящего света (TL-BF) шага ( d ) объединяется с флуоресцентными каналами шага ( a ), что приводит к комбинации положения ламели и флуоресцентного сигнала в одном составном Z- куча. f Чтобы компенсировать наклон ламели, наклон измеряется с помощью сигнала TL-BF, Z-стопка поворачивается соответствующим образом и извлекается Z-срез, соответствующий ламели. г Наконец, извлеченный Z-срез регистрируется на карте крио-ТЕМ с этапа (c ).
e Чтобы объединить информацию о флуоресцентном сигнале из шага ( a ) и положение ламели от шага ( d ), оба Z-стека выравниваются с помощью автоматизированного алгоритма 3D-регистрации. После регистрации изображения канал светлого поля проходящего света (TL-BF) шага ( d ) объединяется с флуоресцентными каналами шага ( a ), что приводит к комбинации положения ламели и флуоресцентного сигнала в одном составном Z- куча. f Чтобы компенсировать наклон ламели, наклон измеряется с помощью сигнала TL-BF, Z-стопка поворачивается соответствующим образом и извлекается Z-срез, соответствующий ламели. г Наконец, извлеченный Z-срез регистрируется на карте крио-ТЕМ с этапа (c ).
Полноразмерное изображение
В качестве доказательства принципа мы первоначально применили рабочий процесс крио-CLEM с посткорреляцией на пластинках для локализации флуоресцентно меченных липидных капель (LD) в клетках A549, поскольку LD легко различимы с помощью крио-ЭМ. Мы могли подтвердить все LD ( n = 11), идентифицированные с помощью крио-ЭМ на 4 криопластинах, с помощью нашего корреляционного подхода (дополнительный рисунок 2) со средней точностью корреляции X – Y 124 нм (стандартное отклонение (SD) = 41 нм, n = 11). Затем мы проанализировали влияние коррекции наклона и извлечения одного среза X–Y нашего рабочего процесса. С этой целью мы повторили корреляцию четырех ламелей, но вместо извлеченного X–Y среза флуоресцентной карты использовали проекцию максимальной интенсивности (MIP). Сравнение обеих корреляций (дополнительный рисунок 3) показало, что коррекция наклона и извлечение одного среза X – Y необходимы для рабочего процесса и уменьшают сигнал вне ламели на 66%. Корреляция с использованием флуоресцентной карты MIP показала 35 сигналов LD вне ламеллы, которые соответствуют LD, удаленным во время измельчения крио-FIB. Путем коррекции наклона и извлечения одного среза X-Y мы смогли уменьшить количество сигналов вне ламеллы с 35 до 12.
Мы могли подтвердить все LD ( n = 11), идентифицированные с помощью крио-ЭМ на 4 криопластинах, с помощью нашего корреляционного подхода (дополнительный рисунок 2) со средней точностью корреляции X – Y 124 нм (стандартное отклонение (SD) = 41 нм, n = 11). Затем мы проанализировали влияние коррекции наклона и извлечения одного среза X–Y нашего рабочего процесса. С этой целью мы повторили корреляцию четырех ламелей, но вместо извлеченного X–Y среза флуоресцентной карты использовали проекцию максимальной интенсивности (MIP). Сравнение обеих корреляций (дополнительный рисунок 3) показало, что коррекция наклона и извлечение одного среза X – Y необходимы для рабочего процесса и уменьшают сигнал вне ламели на 66%. Корреляция с использованием флуоресцентной карты MIP показала 35 сигналов LD вне ламеллы, которые соответствуют LD, удаленным во время измельчения крио-FIB. Путем коррекции наклона и извлечения одного среза X-Y мы смогли уменьшить количество сигналов вне ламеллы с 35 до 12.
Рабочий процесс крио-CLEM после корреляции на пластинках показывает, что ABCA3-eGFP преимущественно коррелирует с пластинчатыми мембранными органеллами
Для локализации LB в клетках A549 мы использовали сверхэкспрессию ABCA3-eGFP, которая индуцирует образование LB-подобных органелл 16 . Через 48 ч после трансфекции клетки A549 содержали в среднем 127 крупных сферических структур на клетку (SD = 50, диапазон: 45–179). Сигнал ABCA3-eGFP преимущественно локализовался на ограничивающей мембране и имел средний диаметр 1,2 мкм (стандартное отклонение = 0,8 мкм, диапазон: 0,1–4,5 мкм), что было выявлено с помощью конфокальной микроскопии (дополнительный рисунок 4). Эти измерения соответствуют диаметру LB, о котором сообщалось ранее (диапазон: 0,1–2,4 мкм), основанному на ЭМ-исследованиях, проведенных на клетках легких 9.0324 1 . Мы временно трансфицировали клетки A549, выращенные на золотых ЭМ-сетках, и дополнительно окрашивали их ядерным красителем и нейтральным липидным красителем, помечающим LD, с намерением использовать их в качестве реперных маркеров, тем самым повышая точность корреляции. Однако сверхэкспрессия ABCA3-eGFP приводит к истощению LD в клетках. Кроме того, нейтральный липидный краситель также локализован в ядре ABCA3-eGFP-позитивных органелл (дополнительный рисунок 5), что не позволяет нам использовать их в качестве реперных маркеров. После витрификации путем глубокой заморозки образец был перенесен при криогенных температурах в крио-LM для картирования сетки ЭМ. Z-стеки в светлом поле проходящего света (TL-BF) и флуоресцентные каналы были получены, покрывая большую площадь (приблизительно 1,2 мм × 1,2 мм) сетки, чтобы максимизировать количество потенциальных областей для последующего фрезерования. Чтобы улучшить контраст и уменьшить сигнал рассеянного света установки широкопольного микроскопа, а также повысить точность корреляции, мы выполнили деконволюцию данных крио-LM (дополнительный рисунок 6). После деконволюции мы смогли различить близко расположенные ABCA3-eGFP-положительные органеллы, которые до деконволюции выглядели как одна органелла (дополнительная рис.
Однако сверхэкспрессия ABCA3-eGFP приводит к истощению LD в клетках. Кроме того, нейтральный липидный краситель также локализован в ядре ABCA3-eGFP-позитивных органелл (дополнительный рисунок 5), что не позволяет нам использовать их в качестве реперных маркеров. После витрификации путем глубокой заморозки образец был перенесен при криогенных температурах в крио-LM для картирования сетки ЭМ. Z-стеки в светлом поле проходящего света (TL-BF) и флуоресцентные каналы были получены, покрывая большую площадь (приблизительно 1,2 мм × 1,2 мм) сетки, чтобы максимизировать количество потенциальных областей для последующего фрезерования. Чтобы улучшить контраст и уменьшить сигнал рассеянного света установки широкопольного микроскопа, а также повысить точность корреляции, мы выполнили деконволюцию данных крио-LM (дополнительный рисунок 6). После деконволюции мы смогли различить близко расположенные ABCA3-eGFP-положительные органеллы, которые до деконволюции выглядели как одна органелла (дополнительная рис. 6d – j). MIP и сшивание данных крио-LM использовались для создания карты, из которой для измельчения были выбраны подходящие области, демонстрирующие большие ABCA3-eGFP-положительные органеллы (рис. 2a). В большинстве случаев надрезы для снятия напряжения, расположенные рядом с криопластинами, использовались для уменьшения изгиба криоламелей перед окончательным этапом фрезерования 9.0324 32 . Сетки, содержащие от четырех до шести самоподдерживающихся ламелей (рис. 2b), были перенесены в крио-ПЭМ, где они были нанесены на карту, чтобы оценить общее качество ламелей и локализовать связанные с мембраной органеллы (рис. 2c). Среди часто присутствующих органелл, таких как митохондрии и ядра, мембраносвязанные структуры с ламеллярной мембранной архитектурой были выбраны для крио-ЭТ в качестве предполагаемых LB. Чтобы получить флуоресцентную карту размолотых клеток для посткорреляции на ламеллах, сетки были нанесены на карту во второй раз с помощью крио-LM после извлечения из крио-TEM (рис.
6d – j). MIP и сшивание данных крио-LM использовались для создания карты, из которой для измельчения были выбраны подходящие области, демонстрирующие большие ABCA3-eGFP-положительные органеллы (рис. 2a). В большинстве случаев надрезы для снятия напряжения, расположенные рядом с криопластинами, использовались для уменьшения изгиба криоламелей перед окончательным этапом фрезерования 9.0324 32 . Сетки, содержащие от четырех до шести самоподдерживающихся ламелей (рис. 2b), были перенесены в крио-ПЭМ, где они были нанесены на карту, чтобы оценить общее качество ламелей и локализовать связанные с мембраной органеллы (рис. 2c). Среди часто присутствующих органелл, таких как митохондрии и ядра, мембраносвязанные структуры с ламеллярной мембранной архитектурой были выбраны для крио-ЭТ в качестве предполагаемых LB. Чтобы получить флуоресцентную карту размолотых клеток для посткорреляции на ламеллах, сетки были нанесены на карту во второй раз с помощью крио-LM после извлечения из крио-TEM (рис. 2d). Обработанные участки на сетке не показали какого-либо специфического флуоресцентного сигнала, вероятно, из-за повреждения пучка электронов, тогда как флуоресценция в окружающем теле клетки сохранилась (дополнительный рисунок 1). В некоторых случаях загрязнение льдом, обнаруженное в TL-BF на ламелле, приводило к автофлуоресцентному сигналу во всех флуоресцентных каналах (рис. 2d, звездочка). Флуоресцентные изображения клетки до и после крио-ПЭМ для всех проанализированных ламелей показаны на дополнительном рисунке 1. Для получения флуоресцентного сигнала, соответствующего положению ламели, мы использовали регистрацию трехмерного изображения и жесткое преобразование с использованием одного из каналов флуоресценции после деконволюции для наложения. оба тома изображения (рис. 2e). Потеря флуоресцентного сигнала при использовании срезов для снятия стресса была незначительной и, следовательно, не мешала корреляции. Результирующий комбинированный стек изображений включает флуоресцентный сигнал перед фрезерованием FIB с сигналом TL-BF после фрезерования, который содержит точное положение ламели.
2d). Обработанные участки на сетке не показали какого-либо специфического флуоресцентного сигнала, вероятно, из-за повреждения пучка электронов, тогда как флуоресценция в окружающем теле клетки сохранилась (дополнительный рисунок 1). В некоторых случаях загрязнение льдом, обнаруженное в TL-BF на ламелле, приводило к автофлуоресцентному сигналу во всех флуоресцентных каналах (рис. 2d, звездочка). Флуоресцентные изображения клетки до и после крио-ПЭМ для всех проанализированных ламелей показаны на дополнительном рисунке 1. Для получения флуоресцентного сигнала, соответствующего положению ламели, мы использовали регистрацию трехмерного изображения и жесткое преобразование с использованием одного из каналов флуоресценции после деконволюции для наложения. оба тома изображения (рис. 2e). Потеря флуоресцентного сигнала при использовании срезов для снятия стресса была незначительной и, следовательно, не мешала корреляции. Результирующий комбинированный стек изображений включает флуоресцентный сигнал перед фрезерованием FIB с сигналом TL-BF после фрезерования, который содержит точное положение ламели. Флуоресцентная карта, соответствующая пластинке, была извлечена из комбинированного стека изображений с поправкой на угол фрезерования (рис. 2f) и, наконец, путем извлечения среза, соответствующего плоскости пластинки (рис. 2g). Мы использовали четыре угла пластинки, а также четко идентифицируемые флуоресцентно меченные маркеры, такие как ядерная оболочка, в качестве ориентиров, чтобы сопоставить извлеченный Z-срез с картой крио-ТЕМ (рис. 2h) путем нежесткой трансформации. Из-за потери LD из-за сверхэкспрессии ABCA3 (дополнительный рисунок 5) мы не смогли использовать LD в качестве реперных маркеров, которые могли бы еще больше повысить точность корреляции в X – Y. Подробный просмотр коррелированной карты крио-ТЕМ показал, что сигнал ABCA3-eGFP перекрывается с многослойными структурами (рис. 2i).
Флуоресцентная карта, соответствующая пластинке, была извлечена из комбинированного стека изображений с поправкой на угол фрезерования (рис. 2f) и, наконец, путем извлечения среза, соответствующего плоскости пластинки (рис. 2g). Мы использовали четыре угла пластинки, а также четко идентифицируемые флуоресцентно меченные маркеры, такие как ядерная оболочка, в качестве ориентиров, чтобы сопоставить извлеченный Z-срез с картой крио-ТЕМ (рис. 2h) путем нежесткой трансформации. Из-за потери LD из-за сверхэкспрессии ABCA3 (дополнительный рисунок 5) мы не смогли использовать LD в качестве реперных маркеров, которые могли бы еще больше повысить точность корреляции в X – Y. Подробный просмотр коррелированной карты крио-ТЕМ показал, что сигнал ABCA3-eGFP перекрывается с многослойными структурами (рис. 2i).
Клетки A549, выращенные на сетках EM, были временно трансфицированы ABCA3-eGFP, а ядра и липидные капли были флуоресцентно помечены перед погружением в заморозку. MIP деконволютивного стека крио-LM, полученного до измельчения FIB. ABCA3-eGFP показан зеленым цветом, а ядра и липидные капли показаны голубым цветом. b Изображение FIB под углом, показывающее одну и ту же область до и после фрезерования соответственно. c Сшитая крио-ТЭМ карта ламели, полученной фрезерованием FIB, полученная при увеличении 8700×. Белые стрелки показывают ориентиры, используемые для корреляции. d MIP стека крио-LM после деконволюции, полученного после визуализации TEM. Флуоресцентный сигнал в теле клетки остается видимым (стрелки). Загрязнение льдом ламеллы (звездочка) показывает аутофлуоресценцию, которая обнаруживается во всех флуоресцентных каналах и, таким образом, не испускается ABCA3-eGFP. e MIP зарегистрированного и выровненного стека изображений. f Срез Z–Y, показывающий наклон ламели, соответствующий углу фрезерования FIB (вверху), и срез Z–Y после поворота Z-образного пакета на 5°. Края ламели, покрытые органометаллической платиной (стрелки), можно увидеть в виде темных точек на Z-Y-срезе.
MIP деконволютивного стека крио-LM, полученного до измельчения FIB. ABCA3-eGFP показан зеленым цветом, а ядра и липидные капли показаны голубым цветом. b Изображение FIB под углом, показывающее одну и ту же область до и после фрезерования соответственно. c Сшитая крио-ТЭМ карта ламели, полученной фрезерованием FIB, полученная при увеличении 8700×. Белые стрелки показывают ориентиры, используемые для корреляции. d MIP стека крио-LM после деконволюции, полученного после визуализации TEM. Флуоресцентный сигнал в теле клетки остается видимым (стрелки). Загрязнение льдом ламеллы (звездочка) показывает аутофлуоресценцию, которая обнаруживается во всех флуоресцентных каналах и, таким образом, не испускается ABCA3-eGFP. e MIP зарегистрированного и выровненного стека изображений. f Срез Z–Y, показывающий наклон ламели, соответствующий углу фрезерования FIB (вверху), и срез Z–Y после поворота Z-образного пакета на 5°. Края ламели, покрытые органометаллической платиной (стрелки), можно увидеть в виде темных точек на Z-Y-срезе. г Извлеченный Z-срез, соответствующий плоскости ламели, с удаленным с помощью вычислений флуоресцентным сигналом за пределами ламели. Белые стрелки указывают ориентиры, наложенные на ориентиры ( c ). h Наложение, показывающее продукт нежесткого выравнивания между картой TEM на панели ( c ) и флуоресцентное изображение на панели ( г ). i Изображение коррелированной ламели с большим увеличением. j Количественная оценка корреляции с мембраносвязанными органеллами. В общей сложности 62 различных коррелированных сигнала eGFP от 6 ламелей были сопоставлены с соответствующими структурами карты крио-ТЕМ. Двадцать восемь сигналов ABCA3-eGFP были отнесены к LB, 19 — к мембраносвязанным органеллам, а 15 не могли быть отнесены к какой-либо структуре. k Количественная оценка точности корреляции. Для 47 успешно коррелированных структур были измерены расстояние и угол до центра соответствующего сигнала ABCA3-eGFP, которые показаны на радиолокационном графике.
г Извлеченный Z-срез, соответствующий плоскости ламели, с удаленным с помощью вычислений флуоресцентным сигналом за пределами ламели. Белые стрелки указывают ориентиры, наложенные на ориентиры ( c ). h Наложение, показывающее продукт нежесткого выравнивания между картой TEM на панели ( c ) и флуоресцентное изображение на панели ( г ). i Изображение коррелированной ламели с большим увеличением. j Количественная оценка корреляции с мембраносвязанными органеллами. В общей сложности 62 различных коррелированных сигнала eGFP от 6 ламелей были сопоставлены с соответствующими структурами карты крио-ТЕМ. Двадцать восемь сигналов ABCA3-eGFP были отнесены к LB, 19 — к мембраносвязанным органеллам, а 15 не могли быть отнесены к какой-либо структуре. k Количественная оценка точности корреляции. Для 47 успешно коррелированных структур были измерены расстояние и угол до центра соответствующего сигнала ABCA3-eGFP, которые показаны на радиолокационном графике. Средняя точность корреляции составляет 450 нм (SD = 249нм). Масштабные линейки: a , d , e , g 20 мкм, c , f , h 5 9 мкм, 9030 мкм
Средняя точность корреляции составляет 450 нм (SD = 249нм). Масштабные линейки: a , d , e , g 20 мкм, c , f , h 5 9 мкм, 9030 мкм
Изображение в полный размер
Мы классифицировали структуры, соответствующие изолированным сигналам ABCA3-eGFP ( n = 62) на шести ламеллах. В результате 45% сигнала ABCA3-eGFP можно отнести к органеллам, содержащим ламеллярные мембраны со смещением менее 1 мкм. В целом 31% сигнала ABCA3-eGFP коррелировало с мембраносвязанными структурами, включая мультивезикулярные тельца (MVB), эндоплазматический ретикулум (ER) или везикулы. Остальные 24% сигнала ABCA3-eGFP не могут быть отнесены к каким-либо связанным с мембраной органеллам, связанным с ABCA3 (рис. 2j, дополнительные данные 1). Коррелированные структуры показали среднее смещение к локальным максимумам соответствующего сигнала ABCA3-eGPF 450 нм (SD = 249нм) (рис. 2k, дополнительные данные 1). Чтобы оценить значимость извлечения только сигнала, соответствующего ламели на этом образце, мы также выполнили корреляцию на основе MIP собранного стопки с поправкой на наклон для трех ламелей. В отличие от извлеченных срезов, 57% сигналов крио-LM не могли быть отнесены к каким-либо связанным с мембраной органеллам, наблюдаемым с помощью крио-ТЕМ, когда для корреляции использовался MIP. Это подтверждает наш анализ LD (дополнительный рисунок 3), показывая, что коррекция наклона и извлечение одного среза X – Y улучшают корреляцию. Таким образом, мы смогли коррелировать 76% сигнала ABCA3-eGFP со средней точностью корреляции X – Y 450 нм с мембраносвязанными органеллами, из которых 45% имели пластинчатую морфологию.
В отличие от извлеченных срезов, 57% сигналов крио-LM не могли быть отнесены к каким-либо связанным с мембраной органеллам, наблюдаемым с помощью крио-ТЕМ, когда для корреляции использовался MIP. Это подтверждает наш анализ LD (дополнительный рисунок 3), показывая, что коррекция наклона и извлечение одного среза X – Y улучшают корреляцию. Таким образом, мы смогли коррелировать 76% сигнала ABCA3-eGFP со средней точностью корреляции X – Y 450 нм с мембраносвязанными органеллами, из которых 45% имели пластинчатую морфологию.
LB содержат плотно упакованные параллельно изогнутые листы мембраны, часто соединенные с ограничивающей мембраной Т-образными соединениями
Мы проанализировали трехмерную архитектуру мембраны на 11 томограммах, содержащих ABCA3-eGFP-положительные органеллы. Все коррелированные органеллы показали внутрипросветные везикулы и плотно упакованные мембранные листы различной кривизны и с регулярным расстоянием в ядре (дополнительная рис. 7). В LB, демонстрирующих большую концентрическую ламеллярную архитектуру (рис. 3, дополнительный фильм 1), мы заметили, что кривизна мембранных листов увеличивается к центру одновременно со сферической трехмерной организацией параллельных листов. Детальный анализ регулярного расстояния между мембранами с помощью быстрого преобразования Фурье (БПФ) центральной области (рис. 3b) выявил три основные частоты, которые соответствуют расстоянию внутри головной группы (3,7 нм -1 ), ширина бислоя (5,6 нм -1 ) и повтор бислоя (11 нм -1 ). Это подтверждается графиком линии плотности той же площади (рис. 3d). Кроме того, мы наблюдали кристаллическую липидную структуру с латеральным повтором 3,4 нм, измеренную с помощью БПФ (рис. 3g).
3, дополнительный фильм 1), мы заметили, что кривизна мембранных листов увеличивается к центру одновременно со сферической трехмерной организацией параллельных листов. Детальный анализ регулярного расстояния между мембранами с помощью быстрого преобразования Фурье (БПФ) центральной области (рис. 3b) выявил три основные частоты, которые соответствуют расстоянию внутри головной группы (3,7 нм -1 ), ширина бислоя (5,6 нм -1 ) и повтор бислоя (11 нм -1 ). Это подтверждается графиком линии плотности той же площади (рис. 3d). Кроме того, мы наблюдали кристаллическую липидную структуру с латеральным повтором 3,4 нм, измеренную с помощью БПФ (рис. 3g).
a Центральный срез реконструированной томограммы, полученной в месте коррелированного сигнала ABCA3-eGFP (корреляция показана на дополнительном рисунке 7k, l). b Детальный вид организации параллельно изогнутой мембраны сердцевины LB. c БПФ-анализ промежутков между мембранами с параллельными изгибами, площадь, используемая для БПФ-анализа, указана в ( a ). Анализ БПФ выявил три основные частоты: 3,7 нм 90 324–1 90 325 (головная группа–головная группа), 5,5 нм 90 324–1 90 325 (ширина двойного слоя) и 11 нм 90 324–1 90 325 (повторение двойного слоя). d График профиля линии, обозначенный пунктирной линией в ( b ), показывает расстояние между мембранами. e Детальный вид кристаллических липидных структур внутри LB. f Параллельные листы наблюдаются в кристаллическом липидном ядре. г FFT-анализ параллельных слоев кристаллической липидной структуры, площадь, используемая для FFT-анализа, указана в ( e ). Анализ БПФ показывает частоту 3,4 -1 нм для интервала. h График профиля линии, обозначенный пунктирной линией в ( f ). Реконструированная томограмма визуализируется как Дополнительный фильм 1.
b Детальный вид организации параллельно изогнутой мембраны сердцевины LB. c БПФ-анализ промежутков между мембранами с параллельными изгибами, площадь, используемая для БПФ-анализа, указана в ( a ). Анализ БПФ выявил три основные частоты: 3,7 нм 90 324–1 90 325 (головная группа–головная группа), 5,5 нм 90 324–1 90 325 (ширина двойного слоя) и 11 нм 90 324–1 90 325 (повторение двойного слоя). d График профиля линии, обозначенный пунктирной линией в ( b ), показывает расстояние между мембранами. e Детальный вид кристаллических липидных структур внутри LB. f Параллельные листы наблюдаются в кристаллическом липидном ядре. г FFT-анализ параллельных слоев кристаллической липидной структуры, площадь, используемая для FFT-анализа, указана в ( e ). Анализ БПФ показывает частоту 3,4 -1 нм для интервала. h График профиля линии, обозначенный пунктирной линией в ( f ). Реконструированная томограмма визуализируется как Дополнительный фильм 1. Масштабные полосы: a 200 нм, e 50 нм, b , f 50 нм.
Масштабные полосы: a 200 нм, e 50 нм, b , f 50 нм.
Изображение в полный размер
Хотя мембранные листы в параллельно-изогнутом расположении чаще всего встречались в СП, спирально закрученные мембранные листы, а также мембраны, образующие замкнутые отсеки, также наблюдались в сердцевине СП (рис. 4, Доп. фильмы 2 и 3). Поскольку клеточные мембраны преимущественно образуют закрытые компартменты, мы затем проанализировали концы параллельных мембранных листов, чтобы получить представление о том, как структурно организован конец бислоя. Часто наблюдалась округлая плотность на концах бислоя, что указывает на то, что ацильные цепи фосфолипидов не подвергаются воздействию водной среды (рис. 4c–e, стрелки). Интересно, что мембранные листы обычно были ориентированы перпендикулярно ограничивающей мембрану LB. Эти листы часто были соединены с пограничной мембраной ЛБ с тонкой плотностью (рис. 4в). Основываясь на перпендикулярной форме соединения, мы называем эти прямые контакты Т-образными соединениями.
a Центральный срез реконструированной томограммы, полученной в месте коррелированного сигнала ABCA3-eGFP (корреляция показана на дополнительных рисунках 7a, b). b Ручная визуализация томограммы ( a ). Ограничивающая мембрана ЛБ отмечена желтым цветом, листы мембраны – зеленым. Для различения отдельных листов мембраны использовались чередующиеся оттенки зеленого цвета. Непрерывные мембраны отмечены красным цветом. Спирально-скрученный мембранный лист помечен оранжевым цветом. c – e Детальные изображения реконструированной томограммы. Перпендикулярно ориентированные мембранные листы по направлению к пограничной мембране часто соединены тонкой плотностью («Т»-соединение) с пограничной мембраной ЛП (стрелки). Листы мембраны имеют округлую плотность на концах мембраны (стрелки). f – h 3D визуализация подробных видов ( c – e ). Эта цифра визуализируется как дополнительные фильмы 2 и 3. Масштабные полосы: a 200 нм, в – д 50 нм.
Эта цифра визуализируется как дополнительные фильмы 2 и 3. Масштабные полосы: a 200 нм, в – д 50 нм.
Изображение с полным размером
Куполообразные белковые комплексы локализованы на пограничной мембране LB
После более тщательного изучения пограничных мембран LB-подобных органелл мы обнаружили куполообразный белковый комплекс, далее именуемый OMDP ( Рис. 5). OMDP казались полыми и были обнаружены вблизи цистерн ЭР, частично украшенных рибосомами (рис. 5a–d). Хотя OMDP встречались редко ( n = 4) в A549, трансфицированном ABCA3-eGFP.клеток, их часто наблюдали ( n = 171) на пограничной мембране LB и MVB в длительной культуре A549 из девяти томограмм (дополнительная рис. 9), а также на пограничной мембране предшественников LB в клетках HSAEpC ( n = 4) (рис. 5e–h). Чтобы получить представление о структуре OMDP, мы вручную извлекли субтомограммы ( n = 116), содержащие OMDP из двух томограмм, и выполнили усреднение субтомограмм (STA) в Dynamo 33 .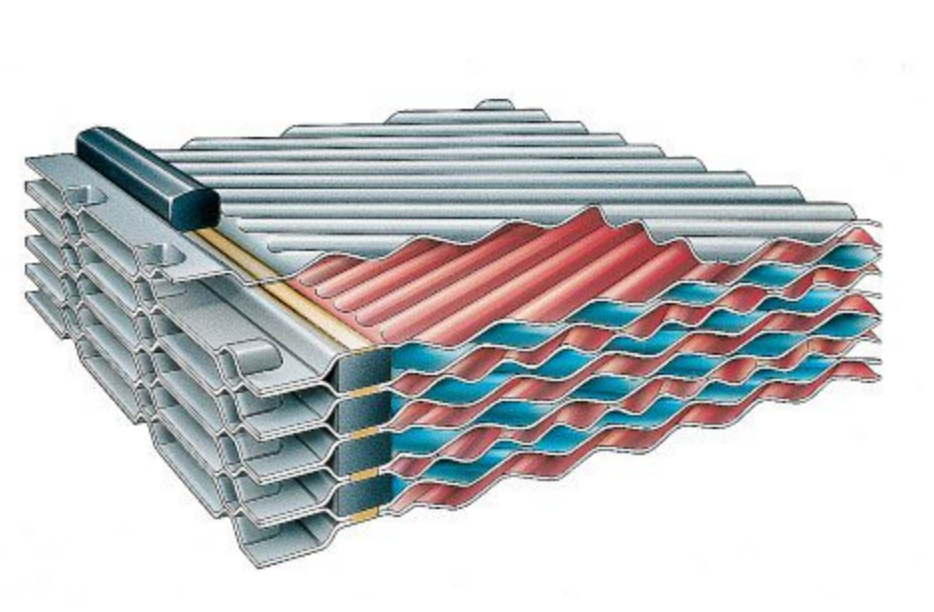 STA OMDP выявил полую куполообразную структуру (рис. 5i–l) с диаметром основания 30 нм, высотой 22 нм с изломом на 7 нм и диаметром вершины 16 нм. Поиск симметрии, выполненный по рассчитанному среднему показателю, показал, что OMDP имеет C 8 симметрия.
STA OMDP выявил полую куполообразную структуру (рис. 5i–l) с диаметром основания 30 нм, высотой 22 нм с изломом на 7 нм и диаметром вершины 16 нм. Поиск симметрии, выполненный по рассчитанному среднему показателю, показал, что OMDP имеет C 8 симметрия.
a Коррелированное изображение пластинки с сигналом ABCA3-eGFP, перекрывающимся с предшественниками MVB и LB. Ядра и митохондрии (мито) не показывают сигнал ABCA3-eGFP. b Срез томограммы, соответствующий области, указанной в ( a ). Указана цистерна ER. Рибосомы отмечены стрелками. c , d Два разных среза одной и той же томограммы, соответствующие области, указанной в ( b ). Сборка OMDP на ограничивающей мембране LB, обращенной к мембране соседней мембраны ER. Всего на томограмме, показанной на ( b ), присутствуют три идентичных сборки. e Срез томограммы, показывающий предшественник LB, содержащий параллельные листы мембраны, соединенные с пограничной мембраной (стрелка) в первичных клетках легкого HSAEpC. LB располагается вблизи промежуточных филаментов (IF), липидных капель (LD) и эндоплазматического ретикулума (ER). ОМДП располагается на пограничной мембране ЛБ (штриховой квадрат). f Срез томограммы, показывающий предшественника ЛБ с изогнутыми листами мембраны и ОМДП, расположенным на пограничной мембране ЛБ (штриховые квадраты). g , h Детали томограммы, показывающие OMDP, соответствующие отмеченной области в ( e , f ). i – k Срезы STA из 105 OMDP, обнаруженные на двух томограммах. i представляет XZ-срез через центр OMDP, j показывает XY-срез через центр, а к через верх ОМДП. l Изоповерхность STA после применения симметрии C 8 . Scale bars: a 5 µm, b , e , f 200 nm, c , d , g , h 20 nm, i – k 10 nm , л 10 нм.
e Срез томограммы, показывающий предшественник LB, содержащий параллельные листы мембраны, соединенные с пограничной мембраной (стрелка) в первичных клетках легкого HSAEpC. LB располагается вблизи промежуточных филаментов (IF), липидных капель (LD) и эндоплазматического ретикулума (ER). ОМДП располагается на пограничной мембране ЛБ (штриховой квадрат). f Срез томограммы, показывающий предшественника ЛБ с изогнутыми листами мембраны и ОМДП, расположенным на пограничной мембране ЛБ (штриховые квадраты). g , h Детали томограммы, показывающие OMDP, соответствующие отмеченной области в ( e , f ). i – k Срезы STA из 105 OMDP, обнаруженные на двух томограммах. i представляет XZ-срез через центр OMDP, j показывает XY-срез через центр, а к через верх ОМДП. l Изоповерхность STA после применения симметрии C 8 . Scale bars: a 5 µm, b , e , f 200 nm, c , d , g , h 20 nm, i – k 10 nm , л 10 нм.
Изображение в натуральную величину
В первичных клетках легких показаны органеллы LB с Т-образными соединениями и OMDP
Для подтверждения того, что архитектура LB, индуцированная экспрессией ABCA3-eGFP в A549представляет собой архитектуру физиологически сформированных LBs, мы использовали первичные эпителиальные клетки малых дыхательных путей человека (HSAEpC), выделенные из дистальной части дыхательных путей человека, а также включили долгосрочные культуры A549, которые, как ранее было показано, восстанавливают дифференцированный фенотип с усиленное производство LB, обнаруженное в первичных клетках AEC2 34 . Крио-ЭТ in situ измельченного HSAEpC выявил пластинчатые органеллы (рис. 5e–h, дополнительная рис. 8) на десяти проанализированных томограммах, соответствующих LB, наблюдаемым в A549.клетки, сверхэкспрессирующие ABCA3. Важно отметить, что LB, обнаруженные в HSAEpC, содержали мембранные листы, прикрепленные к ограничивающей LB мембране через Т-образные соединения (рис. 5e), а изогнутые ламеллярные мембраны были менее конденсированы (рис. 5f). LB в клетках HSAEpC часто обнаруживали вблизи промежуточных филаментов (рис. 5e). Кроме того, цельноклеточная крио-ЭТ долгосрочных клеток A549 также выявила LB (дополнительный рисунок 9) на 6 из 21 проанализированных томограмм. Они демонстрировали морфологию, аналогичную LB, продуцируемым сверхэкспрессией ABCA3-eGFP, и имели средний диаметр 0,58 мкм (стандартное отклонение = 0,25 мкм, диапазон: 0,24–1,1 мкм). Соответствует ранее опубликованным данным 34 , мы обнаружили пластинчатые структуры только в 1 из 15 проанализированных томограмм кратковременно нетрансфицированных клеток А549, что указывает на то, что частое расщепление клеток А549 препятствует формированию LB.
5e), а изогнутые ламеллярные мембраны были менее конденсированы (рис. 5f). LB в клетках HSAEpC часто обнаруживали вблизи промежуточных филаментов (рис. 5e). Кроме того, цельноклеточная крио-ЭТ долгосрочных клеток A549 также выявила LB (дополнительный рисунок 9) на 6 из 21 проанализированных томограмм. Они демонстрировали морфологию, аналогичную LB, продуцируемым сверхэкспрессией ABCA3-eGFP, и имели средний диаметр 0,58 мкм (стандартное отклонение = 0,25 мкм, диапазон: 0,24–1,1 мкм). Соответствует ранее опубликованным данным 34 , мы обнаружили пластинчатые структуры только в 1 из 15 проанализированных томограмм кратковременно нетрансфицированных клеток А549, что указывает на то, что частое расщепление клеток А549 препятствует формированию LB.
Обсуждение
В этом исследовании мы представляем рабочий процесс для посткорреляционной крио-CLEM на ламелях и используем его для изучения трехмерной ультраструктуры LB в невозмущенных условиях. Подходы CLEM широко используются с химически фиксированными образцами или образцами, замещенными замороженными замораживанием под высоким давлением, для идентификации структур-мишеней и редких клеточных событий 22 . Однако такие методы неизбежно скомпрометированы артефактами подготовки образцов из-за обезвоживания, последующего окрашивания и разделения образцов. В частности, структура мембран и богатых липидами органелл плохо сохраняется из-за экстракции липидов, происходящей при обезвоживании образцов органическими растворителями. Крио-ЭМ позволяет напрямую визуализировать мембранные структуры, близкие к их естественному состоянию. Корреляционные подходы, такие как крио-CLEM, могут применяться для идентификации молекул в образце стекловидного тела при условии, что образец остается витрифицированным и свободным от льда во время переноса между крио-LM и крио-TEM 23 . До сих пор рабочие процессы крио-CLEM, включающие фрезерование крио-ФИБ, основывались на высокоточной 3D-корреляции для фрезерования на конкретном участке для нацеливания на интересующую область. Публикация Арнольда и соавт. 29 сообщили о подготовке ламелей с трехмерным нацеливанием на основе крио-LM и корреляции крио-FIB/SEM.
Однако такие методы неизбежно скомпрометированы артефактами подготовки образцов из-за обезвоживания, последующего окрашивания и разделения образцов. В частности, структура мембран и богатых липидами органелл плохо сохраняется из-за экстракции липидов, происходящей при обезвоживании образцов органическими растворителями. Крио-ЭМ позволяет напрямую визуализировать мембранные структуры, близкие к их естественному состоянию. Корреляционные подходы, такие как крио-CLEM, могут применяться для идентификации молекул в образце стекловидного тела при условии, что образец остается витрифицированным и свободным от льда во время переноса между крио-LM и крио-TEM 23 . До сих пор рабочие процессы крио-CLEM, включающие фрезерование крио-ФИБ, основывались на высокоточной 3D-корреляции для фрезерования на конкретном участке для нацеливания на интересующую область. Публикация Арнольда и соавт. 29 сообщили о подготовке ламелей с трехмерным нацеливанием на основе крио-LM и корреляции крио-FIB/SEM. Этот рабочий процесс был установлен на взвешенных клетках, смешанных с флуоресцентными шариками, и требовал больше не распределенного криостола для конфокального микроскопа с вращающимся диском. При таком подходе точность корреляции чувствительна к количеству и распределению крупных реперных шариков и дрейфу, возникающему во время фрезерования 29 . Представленный здесь рабочий процесс полезен тем, что он позволяет проверять целевые структуры после измельчения крио-FIB. Сочетание описанного здесь посткорреляционного подхода на пластинчатой крио-CLEM с ранее описанным целенаправленным фрезерованием 29 позволит проверить нацеленность и гарантировать отсутствие несоответствия флуоресцентного сигнала и прогнозируемого положения томограммы из-за дрейфа предметного столика.
Этот рабочий процесс был установлен на взвешенных клетках, смешанных с флуоресцентными шариками, и требовал больше не распределенного криостола для конфокального микроскопа с вращающимся диском. При таком подходе точность корреляции чувствительна к количеству и распределению крупных реперных шариков и дрейфу, возникающему во время фрезерования 29 . Представленный здесь рабочий процесс полезен тем, что он позволяет проверять целевые структуры после измельчения крио-FIB. Сочетание описанного здесь посткорреляционного подхода на пластинчатой крио-CLEM с ранее описанным целенаправленным фрезерованием 29 позволит проверить нацеленность и гарантировать отсутствие несоответствия флуоресцентного сигнала и прогнозируемого положения томограммы из-за дрейфа предметного столика.
Недавно был разработан прототип микроскопа с крио-LM, интегрированным в камеру крио-FIB микроскопа 35 . Это позволяет контролировать флуоресцентный сигнал во время фрезерования и на готовой ламели. Затем этот сигнал можно напрямую коррелировать без переноса образца.
Затем этот сигнал можно напрямую коррелировать без переноса образца.
В этом исследовании мы реализовали альтернативную стратегию и разработали надежный метод крио-CLEM на пластинках, используя преимущества посткорреляции. Он подходит для прилипших клеток, культивируемых непосредственно на ЭМ-сетках без введения реперных шариков с использованием имеющегося в продаже Leica cryo-LM. Хотя серии наклона (TS) не могут быть получены на основе априорно полученной информации о флуоресценции, посткорреляционный подход дает возможность сопоставлять только высококачественные ламеллы стекловидного тела, оцененные с помощью крио-ПЭМ. Большинство наших попыток выполнить крио-ЛМ после фрезерования, но до крио-ТЭМ, приводили к повышенному загрязнению пластин льдом, что препятствовало высококачественной крио-ЭТ. Кроме того, мы редко обнаруживали флуоресценцию непосредственно на ламеллах, что могло быть связано с пределами чувствительности широкопольного крио-LM Leica.
Хотя деконволюция широкопольной крио-LM успешно использовалась для визуализации тканей 36 , деконволюция не была полностью применена в крио-LM витрифицированных образцов для приложений крио-CLEM. Мы показываем, что деконволюция широкопольных данных крио-LM является необходимым шагом для успешной корреляции и, следовательно, должна применяться также к другим стратегиям крио-CLEM. Вычислительная коррекция наклона и извлечение срезов имеют решающее значение для рабочего процесса даже для крупных органелл, таких как LB. При совместном применении количественная оценка показала, что 76% сигнала ABCA3-eGFP локализовано в связанных с мембраной структурах и органеллах LB. Оставшиеся 24% сигналов не могут быть отнесены к каким-либо структурам, подобным MVB/LB, вероятно, из-за ограниченного осевого разрешения широкопольной микроскопии. Несмотря на то, что деконволюция может улучшить как латеральное, так и аксиальное разрешение, она остается сильно анизотропной и намного превышает толщину ламелей 150–200 нм. Точность корреляции описанной установки микроскопа ограничена аберрациями и числовой апертурой (NA) 0,9объектива с 50-кратным увеличением, используемого в крио-LM, что, в частности, влияет на разрешение в направлении Z.
Мы показываем, что деконволюция широкопольных данных крио-LM является необходимым шагом для успешной корреляции и, следовательно, должна применяться также к другим стратегиям крио-CLEM. Вычислительная коррекция наклона и извлечение срезов имеют решающее значение для рабочего процесса даже для крупных органелл, таких как LB. При совместном применении количественная оценка показала, что 76% сигнала ABCA3-eGFP локализовано в связанных с мембраной структурах и органеллах LB. Оставшиеся 24% сигналов не могут быть отнесены к каким-либо структурам, подобным MVB/LB, вероятно, из-за ограниченного осевого разрешения широкопольной микроскопии. Несмотря на то, что деконволюция может улучшить как латеральное, так и аксиальное разрешение, она остается сильно анизотропной и намного превышает толщину ламелей 150–200 нм. Точность корреляции описанной установки микроскопа ограничена аберрациями и числовой апертурой (NA) 0,9объектива с 50-кратным увеличением, используемого в крио-LM, что, в частности, влияет на разрешение в направлении Z. Разработка линз с улучшенной коррекцией аберраций, криоиммерсионных линз с более высокой числовой апертурой 37 и внедрение конфокальной крио-ЛМ или крио-ЛМ со сверхвысоким разрешением может дополнительно улучшить Z-разрешение 38 и тем самым способствовать более точному удалению внепластинчатого флуоресцентного сигнала в посткорреляционном рабочем процессе. Сверхэкспрессия ABCA3-eGFP привела к меньшему количеству LD по сравнению с A549 без сверхэкспрессии.ячейки (дополнительный рис. 5). Возможно, что сверхэкспрессия ABCA3 приводит к мобилизации нейтральных липидов в LD для синтеза фосфолипидов. Таким образом, мы не могли использовать LD в качестве маркеров корреляции, и окончательная корреляция была выполнена с использованием углов ламели, что ограничивало точность корреляции X – Y до 450 нм (рис. 2k). Для сравнения, мы смогли сопоставить флуоресцентно меченные LD с точностью X – Y 124 нм. Из-за характерной сферической морфологии LD и их повышенной электронной плотности, распознаваемой крио-ЭМ, LD можно использовать в качестве реперных маркеров для повышения точности корреляции.
Разработка линз с улучшенной коррекцией аберраций, криоиммерсионных линз с более высокой числовой апертурой 37 и внедрение конфокальной крио-ЛМ или крио-ЛМ со сверхвысоким разрешением может дополнительно улучшить Z-разрешение 38 и тем самым способствовать более точному удалению внепластинчатого флуоресцентного сигнала в посткорреляционном рабочем процессе. Сверхэкспрессия ABCA3-eGFP привела к меньшему количеству LD по сравнению с A549 без сверхэкспрессии.ячейки (дополнительный рис. 5). Возможно, что сверхэкспрессия ABCA3 приводит к мобилизации нейтральных липидов в LD для синтеза фосфолипидов. Таким образом, мы не могли использовать LD в качестве маркеров корреляции, и окончательная корреляция была выполнена с использованием углов ламели, что ограничивало точность корреляции X – Y до 450 нм (рис. 2k). Для сравнения, мы смогли сопоставить флуоресцентно меченные LD с точностью X – Y 124 нм. Из-за характерной сферической морфологии LD и их повышенной электронной плотности, распознаваемой крио-ЭМ, LD можно использовать в качестве реперных маркеров для повышения точности корреляции. Крио-ЛМ, выполняемый при низкой относительной влажности (30–40%), улучшает как данные крио-ЛМ, так и измельчение крио-ФИБ. Таким образом, интеграция изображений крио-ЛМ в камеры крио-ФИБ/СЭМ, что позволяет получать данные флуоресценции до и после измельчения крио-ФИБ 35 , очень полезно, чтобы избежать передачи образцов. Ультраструктура
Крио-ЛМ, выполняемый при низкой относительной влажности (30–40%), улучшает как данные крио-ЛМ, так и измельчение крио-ФИБ. Таким образом, интеграция изображений крио-ЛМ в камеры крио-ФИБ/СЭМ, что позволяет получать данные флуоресценции до и после измельчения крио-ФИБ 35 , очень полезно, чтобы избежать передачи образцов. Ультраструктура
LB уже давно является предметом споров. Как параллельные прямые, так и концентрические мембранные листы были предложены в качестве основных структурных компонентов LB, и оба типа были рассмотрены CEMOVIS 18 . Однако артефакты вырезания и отсутствие 3D-данных препятствовали дальнейшему пониманию архитектуры органелл. Детальный анализ расстояния между мембранами выявил повторение 11 нм по сравнению с 7,3 нм, обнаруженным с помощью CEMOVIS в легких крысы 18 . Различие в расстоянии между мембранами может быть объяснено сжатием до 50%, которое происходит во время срезов 19 , различиями в стадиях созревания LB или различиями между LB человека и крысы. Тот факт, что первичные клетки легкого человека показали менее компактную упаковку ламеллярных мембран, чем клетки A549, сверхэкспрессирующие ABCA3, указывает на то, что расстояние между мембранами LB является вариабельным, и созревание LB может зависеть от уровней ABCA3 и его регуляции в первичной или иммортализованной клеточной линии.
Тот факт, что первичные клетки легкого человека показали менее компактную упаковку ламеллярных мембран, чем клетки A549, сверхэкспрессирующие ABCA3, указывает на то, что расстояние между мембранами LB является вариабельным, и созревание LB может зависеть от уровней ABCA3 и его регуляции в первичной или иммортализованной клеточной линии.
Пост-корреляционная крио-CLEM на ламелях в сочетании с крио-ЭТ in situ положительных органелл ABCA3-eGFP позволила нам раскрыть архитектуру LB в 3D. Ядро имеет несколько концентрических листов мембраны с увеличивающейся кривизной к центру. Однако ни одна из наблюдаемых СП не была точно центросимметричной, что было предложено ранее на основе поляризованного LM 39 . Т-образные соединения можно наблюдать на внутреннем листке пограничной мембраны, где мембраны вдавливаются в липидное ядро в виде параллельных слоев, что соответствует модели, предложенной Пересом-Гилом 9.0324 17 . В дополнение к параллельно изогнутым листам мембраны мы иногда наблюдали кристаллические липидные структуры внутри ядра LB с регулярным интервалом 3,4 нм -1 , как показало БПФ. Это расстояние соответствует кристаллам эфира холестерина, обнаруженным в LD 40 .
Это расстояние соответствует кристаллам эфира холестерина, обнаруженным в LD 40 .
Наше обнаружение того, что сигнал ABCA3-eGFP локализуется не только в LB, но и в других связанных с мембраной органеллах, таких как MVB, ER или везикулы, совпадает с предыдущими данными о переносе и протеолитическом процессинге ABCA3 в LAMP3-позитивных везикулах 41 . Однако распределение ABCA3 и его фракция на LB могут различаться в клетках A549 и первичных клетках легкого. Поскольку LB, обнаруженные в клетках A549, экспрессирующих ABCA3-eGFP, содержат не только ламеллярные мембраны, но также кристаллические липидные структуры и везикулы, эти органеллы скорее представляют собой «композитные тельца», форму предшественника LB. LB, обнаруженные в первичных клетках легкого, были морфологически сходны с таковыми, индуцированными сверхэкспрессией ABCA3-eGFP, что указывает на то, что ни ламеллярная организация, ни «T»-соединения, обнаруженные в ABCA3-eGFP-позитивных органеллах, не являются артефактами сверхэкспрессии. Примечательно, что все LB-подобные органеллы, обнаруженные в HSAEpC, показали морфологию ранних предшественников LB, и мы не наблюдали зрелых LB, обнаруженных в долгосрочной культуре A549.ячейки (дополнительный рис. 9). Стоит отметить, что HSAEpC представляют собой смесь типов эпителиальных клеток, и только часть из них представляет собой клетки ACE2 42 . Долгосрочная культура A549 показала повышенное количество органелл по сравнению с краткосрочными нетрансфицированными клетками. Поскольку маркер LB не использовался для локализации LB в долгосрочной культуре A549, мы провели крио-ET как на крио-FIB-размолотых клетках, так и на периферии целых клеток. Интересно, что большинство крупных LB были обнаружены на периферии клеток A549.длительная культура, при которой происходит созревание ЛБ и секреция сурфактанта. Посткорреляционный рабочий процесс крио-CLEM на ламеллах может применяться для изучения локализации других белков, участвующих в созревании LB, таких как Rab38, который играет незаменимую роль в поддержании морфологии LB 43 .
Примечательно, что все LB-подобные органеллы, обнаруженные в HSAEpC, показали морфологию ранних предшественников LB, и мы не наблюдали зрелых LB, обнаруженных в долгосрочной культуре A549.ячейки (дополнительный рис. 9). Стоит отметить, что HSAEpC представляют собой смесь типов эпителиальных клеток, и только часть из них представляет собой клетки ACE2 42 . Долгосрочная культура A549 показала повышенное количество органелл по сравнению с краткосрочными нетрансфицированными клетками. Поскольку маркер LB не использовался для локализации LB в долгосрочной культуре A549, мы провели крио-ET как на крио-FIB-размолотых клетках, так и на периферии целых клеток. Интересно, что большинство крупных LB были обнаружены на периферии клеток A549.длительная культура, при которой происходит созревание ЛБ и секреция сурфактанта. Посткорреляционный рабочий процесс крио-CLEM на ламеллах может применяться для изучения локализации других белков, участвующих в созревании LB, таких как Rab38, который играет незаменимую роль в поддержании морфологии LB 43 .
Мы идентифицировали куполообразный белковый комплекс на внешней мембране LB как в клетках A549, так и в первичных клетках легкого, о котором, насколько нам известно, еще не сообщалось. STA выявил, что OMDP имеют полую клеткообразную структуру, однако ограниченное количество усредненных OMDP не позволило получить молекулярные детали комплекса. Тем не менее, исходя из указанного C 8 симметрии мы предполагаем, что комплекс построен из множества субъединиц, состоящих из белковых сборок, симметрично организованных вокруг одной оси вращения. Примечательно, что мы часто наблюдали структурно хорошо охарактеризованный белок главного свода (MVP) 13 MDa в цитозоле клеток A549, который образует большие каркасные структуры с 39-кратной диэдрической симметрией 44 . Однако размеры концевых колпачков MVP отличаются от OMDP, что указывает на то, что OMDP не является производным от MVP (дополнительный рис. 9).). Хотя OMDP редко обнаруживались в клетках, экспрессирующих ABCA3-eGFP, они часто наблюдались в долговременной культуре A549.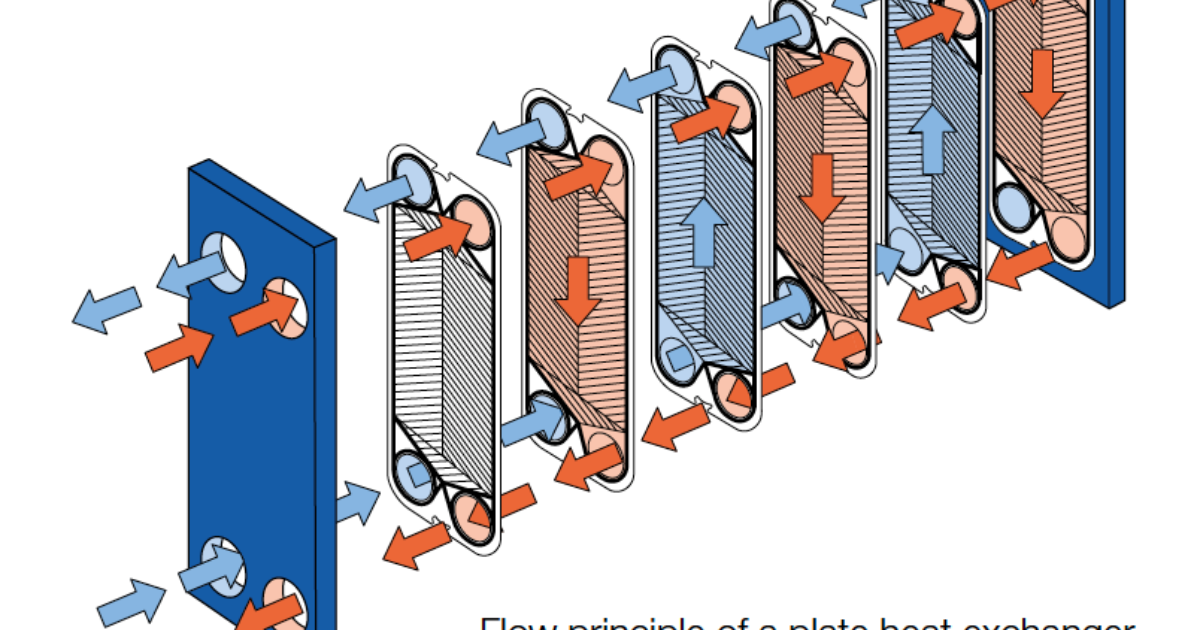 Мы полагаем, что низкая встречаемость OMDP в клетках A549 со сверхэкспрессией ABAC3-eGFP может быть объяснена высокой занятостью ABCA3 на ограничивающей мембране, вызывающей вытеснение других белков. Поскольку OMDP также наблюдались на MVB, мы предполагаем, что этот комплекс играет более общую роль в эндолизосомном транспорте. Состав OMDP выходит за рамки данного исследования и требует дальнейшего изучения. Поскольку зрелые LB подвергаются экзоцитозу, мы предполагаем, что OMDPs могут быть вовлечены в связывание органелл или в регуляцию слияния между ограничивающей мембраной и плазматической мембраной. OMDP можно наблюдать на криоэлектронной томограмме in situ (EMDB 4604) цианобактерии 9.0324 45 , что указывает на то, что этот комплекс может сохраняться в нескольких областях жизни и может быть задействован в биологии мембран в целом.
Мы полагаем, что низкая встречаемость OMDP в клетках A549 со сверхэкспрессией ABAC3-eGFP может быть объяснена высокой занятостью ABCA3 на ограничивающей мембране, вызывающей вытеснение других белков. Поскольку OMDP также наблюдались на MVB, мы предполагаем, что этот комплекс играет более общую роль в эндолизосомном транспорте. Состав OMDP выходит за рамки данного исследования и требует дальнейшего изучения. Поскольку зрелые LB подвергаются экзоцитозу, мы предполагаем, что OMDPs могут быть вовлечены в связывание органелл или в регуляцию слияния между ограничивающей мембраной и плазматической мембраной. OMDP можно наблюдать на криоэлектронной томограмме in situ (EMDB 4604) цианобактерии 9.0324 45 , что указывает на то, что этот комплекс может сохраняться в нескольких областях жизни и может быть задействован в биологии мембран в целом.
На основании структурного анализа ABCA3-позитивных органелл, а также LB, обнаруженных в долговременной культуре A549, мы предлагаем модель биогенеза LB (рис. 6): ABCA3-опосредованная липидная асимметрия мембраны заставляет внутреннюю створку выталкивать бислои в просвет, образуя Т-образные соединения. Возможно, сам ABCA3 или другие белки ответственны за концентрацию фосфолипидов в листах мембраны. SP-B представляет собой небольшой белок, содержащий амфипатические спирали 9.0324 14 и, таким образом, может отвечать за стабилизацию концов мембранных листов и предотвращение их слияния в везикулы. Эти листы отделяются от ограничивающей мембраны LB и формируются в соответствии с основной сферической кривизной LB, чтобы максимизировать емкость хранения липидов на заданный объем. Наконец, OMDP, наблюдаемый на ограничивающей мембране, может регулировать доставку LB или участвовать в экзоцитозе.
6): ABCA3-опосредованная липидная асимметрия мембраны заставляет внутреннюю створку выталкивать бислои в просвет, образуя Т-образные соединения. Возможно, сам ABCA3 или другие белки ответственны за концентрацию фосфолипидов в листах мембраны. SP-B представляет собой небольшой белок, содержащий амфипатические спирали 9.0324 14 и, таким образом, может отвечать за стабилизацию концов мембранных листов и предотвращение их слияния в везикулы. Эти листы отделяются от ограничивающей мембраны LB и формируются в соответствии с основной сферической кривизной LB, чтобы максимизировать емкость хранения липидов на заданный объем. Наконец, OMDP, наблюдаемый на ограничивающей мембране, может регулировать доставку LB или участвовать в экзоцитозе.
Эта модель биогенеза LB основана на наблюдаемых ультраструктурных особенностях LB с помощью крио-ПЭМ-анализа. На всех стадиях биогенеза ЛБ ОМДП присутствуют на лимитирующей мембране ЛБ. a Ранняя стадия: ABCA3 переворачивает фосфолипиды с листка внешней мембраны на внутренний, что приводит к асимметрии фосфолипидов по отношению к внутреннему листку. Это приводит к образованию мембранных листов, которые напрямую связаны с пограничной мембраной Т-образными соединениями. b Промежуточная стадия: листы мембраны растут дальше и в конечном итоге отделяются от ограничивающей мембраны. Отдельные мембранные листы начинают складываться в изогнутые формы. c Заключительная стадия: полость заполнена листами мембран, которые образуют комбинацию различных структурных структур, включая спиральные, параллельные прямые и изогнутые мембраны, а также кристаллические липидные структуры.
a Ранняя стадия: ABCA3 переворачивает фосфолипиды с листка внешней мембраны на внутренний, что приводит к асимметрии фосфолипидов по отношению к внутреннему листку. Это приводит к образованию мембранных листов, которые напрямую связаны с пограничной мембраной Т-образными соединениями. b Промежуточная стадия: листы мембраны растут дальше и в конечном итоге отделяются от ограничивающей мембраны. Отдельные мембранные листы начинают складываться в изогнутые формы. c Заключительная стадия: полость заполнена листами мембран, которые образуют комбинацию различных структурных структур, включая спиральные, параллельные прямые и изогнутые мембраны, а также кристаллические липидные структуры.
Изображение в полный размер
В заключение мы разработали и внедрили надежный рабочий процесс крио-CLEM с посткорреляцией на пластинках, оптимизированный для прикрепленных эпителиальных клеток человека, который совместим с установками крио-LM с широким полем. Его можно применять для подтверждения идентичности клеточных органелл с использованием четко определенного флуоресцентного маркера для выявления структурных деталей органелл в контексте нативной клеточной среды.
Методы
Клеточные линии и клеточные культуры
Клетки A549 31 были приобретены в Европейской коллекции аутентифицированных клеточных культур ECACC (лот 16J012) и культивированы в среде DMEM/F12 (ThermoFisher Scientific) с добавлением 10% (об./об.) эмбриональной бычьей сыворотки (FBS) (ThermoFisher Scientific) и 100 ЕД/мл пенициллина-стрептомицина (ThermoFisher Scientific) при 37 °C, 5% CO 2 . Первичные эпителиальные клетки малых дыхательных путей человека HSAEpC были приобретены у PromoCell (лот 414Z012.2) и использованы в пассаже 3. Клетки HSAEcP поддерживали в базальной среде эпителиальных клеток малых дыхательных путей (PromoCell, C-21275) с добавлением среды для роста эпителиальных клеток малых дыхательных путей. Смесь добавок (PromoCell, C-39175) и 100 ЕД/мл пенициллина-стрептомицина (ThermoFisher Scientific) в течение 4 недель (среда заменялась 3 раза в неделю), а для отделения клеток от чашки для культивирования перед посевом использовали набор Detach Kit (PromoCell, C-41210). ячейки на ЭМ сетках. Долговременную культуру A549 поддерживали без пересева в соответствии с протоколом Cooper et al. 34 в Ham’s F12 (Sigma-Aldrich) с добавлением 10% FBS и 100 МЕ/мл пенициллина-стрептомицина (ThermoFisher Scientific) в течение 28 дней. Среду заменяли два раза в неделю. Для подготовки образцов к глубокой заморозке, 200 меш Quantifoil 9.Сетки 0324™ Au R2/2 подвергали плазменной очистке в Gatan Solarus 950 (Gatan) в течение 10 с, стерилизовали погружением в 70% этанол, помещали в чашку диаметром 35 мм (ThermoFischer Scientific), покрытую тонким слоем полидиметилсилоксана ( PDMS, Dow Europe GmbH) и инкубировали в 1 мл среды при 37°С в течение 30 мин. Затем среду удаляли и 1,2 × 10 5 клеток высевали на сетки, помещенные в чашку, покрытую PDMS, содержащую 2 мл среды. На следующий день клетки временно трансфицировали 3 мкг плазмиды pE-hABCA3-eGFP (экспрессия hABCA3-eGFP контролируется промотором CMV) в реагенте для трансфекции Trans-IT LT1 (Mirus).
ячейки на ЭМ сетках. Долговременную культуру A549 поддерживали без пересева в соответствии с протоколом Cooper et al. 34 в Ham’s F12 (Sigma-Aldrich) с добавлением 10% FBS и 100 МЕ/мл пенициллина-стрептомицина (ThermoFisher Scientific) в течение 28 дней. Среду заменяли два раза в неделю. Для подготовки образцов к глубокой заморозке, 200 меш Quantifoil 9.Сетки 0324™ Au R2/2 подвергали плазменной очистке в Gatan Solarus 950 (Gatan) в течение 10 с, стерилизовали погружением в 70% этанол, помещали в чашку диаметром 35 мм (ThermoFischer Scientific), покрытую тонким слоем полидиметилсилоксана ( PDMS, Dow Europe GmbH) и инкубировали в 1 мл среды при 37°С в течение 30 мин. Затем среду удаляли и 1,2 × 10 5 клеток высевали на сетки, помещенные в чашку, покрытую PDMS, содержащую 2 мл среды. На следующий день клетки временно трансфицировали 3 мкг плазмиды pE-hABCA3-eGFP (экспрессия hABCA3-eGFP контролируется промотором CMV) в реагенте для трансфекции Trans-IT LT1 (Mirus). Через 48 ч после трансфекции среду удаляли и клетки инкубировали в свежей среде, содержащей 1 мкг/мл Hoechst 33342 (Sigma-Aldrich) и 150 нМ LipiBlue (Dojindo Europe) для окрашивания ядер и LD соответственно. После 30 мин инкубации клетки трижды промывали средой.
Через 48 ч после трансфекции среду удаляли и клетки инкубировали в свежей среде, содержащей 1 мкг/мл Hoechst 33342 (Sigma-Aldrich) и 150 нМ LipiBlue (Dojindo Europe) для окрашивания ядер и LD соответственно. После 30 мин инкубации клетки трижды промывали средой.
Витрификация
Клетки подвергали погруженной заморозке в жидкий этан сразу после реперного окрашивания с использованием автоматического погружного морозильника Leica EM GP2. Температура криогена была установлена на -183 ° C, а камера — на 25 ° C и влажность 95%. Сетки промокали с обратной стороны бумагой Whatman ® Type 1 в течение 4–5 с. Всего на сетку было добавлено 5 мкл среды непосредственно перед погружением в заморозку. Для образцов, которые использовались для томографии целых клеток, были добавлены реперные точки Protein-A Gold с номинальным диаметром 10 нм (Aurion). Сетки были вставлены в AutoGrids ™ (ThermoFisher Scientific), предназначенный для фрезерования FIB.
Криофлуоресцентная широкопольная микроскопия
Флуоресцентные карты образцов стекловидного тела были получены с использованием широкопольного микроскопа EM Cryo-CLEM (Leica), оснащенного 50-кратным воздушным объективом (NA 0,9), металлогалогенным источником света (EL6000), воздушно-капельным охлаждаемый детектор (DFC9000GT) и криостол, охлаждаемый до -190 °C в помещении с относительной влажностью от 40 до 60%. Квадрат размером примерно 1,2 мм на 1,2 мм был получен в центре сетки с помощью LAS X Navigator. Для каждого поля зрения вокруг точки автофокусировки был получен симметричный Z-стек размером 30 мкм с интервалом 300 нм. Были использованы следующие каналы: TL-BF, DAPI (пример: BP 350/50, Em: 460/50) и GFP (пример: BP470/50, Em: 525/50). Файлы TIF были экспортированы из LAS X, а карта MIP была сшита с помощью плагина сшивки Grid/Collection 9.0324 46 в ImageJ/FIJI 47 .
Квадрат размером примерно 1,2 мм на 1,2 мм был получен в центре сетки с помощью LAS X Navigator. Для каждого поля зрения вокруг точки автофокусировки был получен симметричный Z-стек размером 30 мкм с интервалом 300 нм. Были использованы следующие каналы: TL-BF, DAPI (пример: BP 350/50, Em: 460/50) и GFP (пример: BP470/50, Em: 525/50). Файлы TIF были экспортированы из LAS X, а карта MIP была сшита с помощью плагина сшивки Grid/Collection 9.0324 46 в ImageJ/FIJI 47 .
Деконволюция данных широкопольной криофлуоресцентной микроскопии
Для повышения четкости флуоресцентного сигнала стопки изображений, содержащие интересующие клетки, были подвергнуты деконволюции в AutoQuant X3 (Media Cybernetics). Теоретическая функция рассеяния точки использовалась на основе спецификаций микроскопа и объектива (числовая апертура 0,9, линза 50×, показатель преломления 1) и уточнялась в течение 100 итераций.
Измельчение Cryo-FIB-SEM
После криофлуоресцентной визуализации были подготовлены самонесущие ламели с использованием крио-FIB фрезерования на двухлучевом микроскопе Aquilos FIB-SEM в помещении с контролируемой относительной влажностью от 30 до 40% (ThermoFisher Scientific), как впервые описано Rigort et al. др. 20 . Трансфицированные клетки отбирали с помощью корреляции со сшитой картой флуоресценции MIP, полученной на предыдущем этапе с использованием программного обеспечения MAPS 3.3 (ThermoFisher Scientific). Клетки покрывали слоем металлоорганической платины на 5–6 с и постепенно утоняли в 4 этапа при угле наклона столика 15–18° с помощью Ga + для получения ламелей толщиной 150–200 нм после окончательного этапа фрезерования. По возможности использовались микрокомпенсаторы, как описано Wolff et al. 32 для повышения стабильности пластин. Прогресс контролировали с помощью СЭМ-изображения при 2,2–10 кВ с детекторами ETD и T1.
др. 20 . Трансфицированные клетки отбирали с помощью корреляции со сшитой картой флуоресценции MIP, полученной на предыдущем этапе с использованием программного обеспечения MAPS 3.3 (ThermoFisher Scientific). Клетки покрывали слоем металлоорганической платины на 5–6 с и постепенно утоняли в 4 этапа при угле наклона столика 15–18° с помощью Ga + для получения ламелей толщиной 150–200 нм после окончательного этапа фрезерования. По возможности использовались микрокомпенсаторы, как описано Wolff et al. 32 для повышения стабильности пластин. Прогресс контролировали с помощью СЭМ-изображения при 2,2–10 кВ с детекторами ETD и T1.
Получение серии Tilt и реконструкция томограммы
После подготовки ламелей сетки были перенесены в крио-ТЕМ Titan Krios, работающий на 300 кВ (ThermoFisher Scientific), оснащенный детектором прямых электронов K3 и визуализирующим фильтром Gatan (Gatan). Получение изображения контролировалось с помощью SerialEM 9. 0324 48 . Сначала были получены карты с малым увеличением при 135-кратном увеличении для обнаружения ламелей, затем были получены карты со средним увеличением (МММ) при 8700-кратном увеличении для корреляции и идентификации участков для получения TS при 26 000-кратном или 33 000-кратном увеличении (соответствующие размеры пикселей). на уровне образца: 3,356 и 2,671 Å соответственно). TS были получены с использованием дозосимметричной схемы 49 с постоянной дозой электронов ок. 3 e − / Å 2 на проекцию, расфокусировка цели −5 мкм, щель энергетического фильтра при 20 эВ, охватывающая диапазон от 60° до −60° с шагом 3°. Томографию на пластинке выполняли с наклоном предметного столика на 6°, чтобы компенсировать предварительный наклон пластинки по отношению к сетке. Каждая проекция была получена в режиме подсчета с использованием фракционирования дозы и 20–40 отдельных кадров, которые были выровнены и суммированы с помощью плагина SEMCCD в SerialEM.
0324 48 . Сначала были получены карты с малым увеличением при 135-кратном увеличении для обнаружения ламелей, затем были получены карты со средним увеличением (МММ) при 8700-кратном увеличении для корреляции и идентификации участков для получения TS при 26 000-кратном или 33 000-кратном увеличении (соответствующие размеры пикселей). на уровне образца: 3,356 и 2,671 Å соответственно). TS были получены с использованием дозосимметричной схемы 49 с постоянной дозой электронов ок. 3 e − / Å 2 на проекцию, расфокусировка цели −5 мкм, щель энергетического фильтра при 20 эВ, охватывающая диапазон от 60° до −60° с шагом 3°. Томографию на пластинке выполняли с наклоном предметного столика на 6°, чтобы компенсировать предварительный наклон пластинки по отношению к сетке. Каждая проекция была получена в режиме подсчета с использованием фракционирования дозы и 20–40 отдельных кадров, которые были выровнены и суммированы с помощью плагина SEMCCD в SerialEM. Томографию целых клеток проводили, как указано выше, за исключением того, что TS был получен при увеличении 33000× (размер пикселя на уровне образца: 4,302 Å) с использованием детектора прямых электронов K2 вместо детектора прямых электронов K3.
Томографию целых клеток проводили, как указано выше, за исключением того, что TS был получен при увеличении 33000× (размер пикселя на уровне образца: 4,302 Å) с использованием детектора прямых электронов K2 вместо детектора прямых электронов K3.
TS обработан с использованием пакета IMOD 50 . TS были выровнены с помощью отслеживания исправлений. Перед реконструкцией передаточная функция контраста была скорректирована путем обращения фазы, а микрофотографии подверглись дозофильтрации. Томограммы были реконструированы с использованием алгоритма взвешенной обратной проекции с SIRT-подобным фильтром, эквивалентным десяти итерациям. Для спектрального анализа мощности на рис. 4 томограммы были реконструированы с использованием алгоритма SIRT, реализованного в 3dmod.
3D визуализация реконструированных томограмм
Визуализации были созданы в Amira 2019.3 (ThermoFisher Scientific). Во-первых, был применен фильтр усиления мембраны. С помощью инструмента сегментации Top Hat был создан первый 3D-рендеринг. На основе этой исходной модели все мембраны были сегментированы вручную.
На основе этой исходной модели все мембраны были сегментированы вручную.
Усреднение субтомограммы
СТА выполнена в Dynamo 33 . Частицы ( n = 116) были выбраны вручную на 2 томограммах, и 100 субтомограмм вокселей были извлечены с использованием функции обрезки на стенке с исходной ориентацией частиц, приписанной нормально к сегментированной ограничивающей мембране LB. Первоначальный эталон был создан путем усреднения всех 105 частиц, а среднее значение было итеративно уточнено с использованием маски в форме полукупола. Сканирование симметрии было выполнено с использованием сфокусированной маски на верхней части узла клетки. Впоследствии среднее значение уточнялось с использованием C 8 симметрия.
Рабочий процесс посткорреляционной корреляции на пластинках
После витрификации флуоресцентно меченных клеток методом глубокой заморозки сетки были нанесены на карту Leica cryo-LM. Сетка была перенесена на двухлучевой крио-FIB/SEM. Путем низкоточной корреляции флуоресцентной карты MIP с картой сетки SEM были выбраны интересующие клетки и измельчены FIB. После второго переноса образца в крио-ПЭМ на 300 кВ были нанесены на карту ламели, выбраны интересующие области и получен TS. Чтобы облегчить высокоточную пост-корреляцию ламелей, сетки были восстановлены после получения крио-ЭТ, а ламели были нанесены на карту во второй раз в крио-ЛМ, чтобы выявить точное положение ламелей на сетке. Чтобы объединить флуоресцентный сигнал с первой карты крио-ЛМ с информацией о положении ламелей на второй карте крио-ЛМ, стеки деконволюции были зарегистрированы в MATLAB 2018b (MathWorks) с использованием функция imregtform с пользовательским скриптом (см. раздел «Доступность кода»). Матрица жесткого преобразования, включающая перевод и вращение, была рассчитана на основе одного и того же флуоресцентного канала обеих карт крио-LM соответственно. Остальные каналы, включая TL-BF из второй карты крио-LM, были преобразованы с применением рассчитанной матрицы преобразования.
Путем низкоточной корреляции флуоресцентной карты MIP с картой сетки SEM были выбраны интересующие клетки и измельчены FIB. После второго переноса образца в крио-ПЭМ на 300 кВ были нанесены на карту ламели, выбраны интересующие области и получен TS. Чтобы облегчить высокоточную пост-корреляцию ламелей, сетки были восстановлены после получения крио-ЭТ, а ламели были нанесены на карту во второй раз в крио-ЛМ, чтобы выявить точное положение ламелей на сетке. Чтобы объединить флуоресцентный сигнал с первой карты крио-ЛМ с информацией о положении ламелей на второй карте крио-ЛМ, стеки деконволюции были зарегистрированы в MATLAB 2018b (MathWorks) с использованием функция imregtform с пользовательским скриптом (см. раздел «Доступность кода»). Матрица жесткого преобразования, включающая перевод и вращение, была рассчитана на основе одного и того же флуоресцентного канала обеих карт крио-LM соответственно. Остальные каналы, включая TL-BF из второй карты крио-LM, были преобразованы с применением рассчитанной матрицы преобразования. Флуоресцентные каналы первой крио-LM карты были объединены с преобразованным каналом TL-BF второй крио-LM в FIJI/ImageJ и сохранены как составной TIF. Чтобы компенсировать угол фрезерования FIB, композитный пакет был повторно нарезан с использованием FIJI/ImageJ. Используя канал TL-BF, был измерен фактический наклон ламелей в пакете, и композитный пакет был повернут соответствующим образом, чтобы компенсировать наклон. Z-срез пластинки был идентифицирован и извлечен после второго повторного среза в FIJI/ImageJ. Этот единственный срез с поправкой на наклон был сопоставлен со сшитой TEM-картой с использованием нежесткого 2D-преобразования в ec-CLEM 9.0324 51 с использованием четырех углов ламеллы, а также четко идентифицируемых флуоресцентно помеченных органелл, таких как LD, в качестве ориентиров.
Флуоресцентные каналы первой крио-LM карты были объединены с преобразованным каналом TL-BF второй крио-LM в FIJI/ImageJ и сохранены как составной TIF. Чтобы компенсировать угол фрезерования FIB, композитный пакет был повторно нарезан с использованием FIJI/ImageJ. Используя канал TL-BF, был измерен фактический наклон ламелей в пакете, и композитный пакет был повернут соответствующим образом, чтобы компенсировать наклон. Z-срез пластинки был идентифицирован и извлечен после второго повторного среза в FIJI/ImageJ. Этот единственный срез с поправкой на наклон был сопоставлен со сшитой TEM-картой с использованием нежесткого 2D-преобразования в ec-CLEM 9.0324 51 с использованием четырех углов ламеллы, а также четко идентифицируемых флуоресцентно помеченных органелл, таких как LD, в качестве ориентиров.
Анализ точности корреляции
Для оценки корреляции и точности X–Y посткорреляционного рабочего процесса крио-CLEM на пластинке была проанализирована окончательная корреляция карты MMM и крио-LM. Локальные максимумы карты крио-ЛМ были рассчитаны с использованием FIJI/ImageJ с использованием порога допуска 10. Для каждого локального максимума ультраструктуры коррелированного МММ в сферической области (радиус = 1 мкм) классифицировались как LD, LB или мембраносвязанные органеллы (MVB, ER и везикула). Митохондриальные или ядерные мембраны, а также участки без каких-либо мембраносвязанных структур классифицировались как неспецифические. Для каждой заданной ультраструктуры были измерены расстояние и угол до центра локального максимума, которые были построены в виде радиолокационного графика для оценки точности корреляции.
Локальные максимумы карты крио-ЛМ были рассчитаны с использованием FIJI/ImageJ с использованием порога допуска 10. Для каждого локального максимума ультраструктуры коррелированного МММ в сферической области (радиус = 1 мкм) классифицировались как LD, LB или мембраносвязанные органеллы (MVB, ER и везикула). Митохондриальные или ядерные мембраны, а также участки без каких-либо мембраносвязанных структур классифицировались как неспецифические. Для каждой заданной ультраструктуры были измерены расстояние и угол до центра локального максимума, которые были построены в виде радиолокационного графика для оценки точности корреляции.
Конфокальная флуоресцентная микроскопия
Конфокальная микроскопия клеток A549, трансфицированных ABCA3-eGFP, проводилась с использованием лазерного сканирующего конфокального микроскопа SP8 TCS (Leica), оснащенного масляным иммерсионным объективом с числовой апертурой 63×1,4 и камерой контроля окружающей среды, нагретой до 37 °C. . Анализ изображения выполняли в программе ImageJ/FIJI 47 . Автоматизированная 3D-сегментация структур ABCA3-eGFP была выполнена в Imaris 9.5.1 (Bitplane) после деконволюции в AutoQuant X3.1 (Media Cybernetics).
Автоматизированная 3D-сегментация структур ABCA3-eGFP была выполнена в Imaris 9.5.1 (Bitplane) после деконволюции в AutoQuant X3.1 (Media Cybernetics).
Статистика и воспроизводимость
Рабочий процесс был оптимизирован в ходе нескольких испытаний. Представленные данные получены из двух независимых биологических повторов. Оценка размера выборки не проводилась. Структуры, соответствующие изолированным сигналам ABCA3-eGFP ( n = 62), классифицировали на шести ламеллах. Доступны исходные данные, содержащие измерения и количественные оценки (дополнительные данные 1).
Сводка отчета
Дополнительную информацию о дизайне исследования можно найти в Сводке отчета об исследовании природы, связанной с этой статьей.
Доступность данных
Данные EM предоставляются в базе данных EMDB под номерами доступа: EMD-10818, EMD-10819, EMD-10820, EMD-10821, EMD-11929 и EMD-11931. Исходные данные приводятся вместе с настоящей статьей.
Доступность кода
Макрос FIJI/ImageJ для сборки флуоресцентной карты и код MATLAB, используемый для регистрации 3D-стека, доступен на GitHub по адресу https://github. com/chlanda-lab.
com/chlanda-lab.
Ссылки
Schmitz, G. & Muller, G. Структура и функция пластинчатых тел, липидно-белковые комплексы, участвующие в хранении и секреции клеточных липидов. J. Lipid Res. 32 , 1539–1570 (1991).
КАС пабмед Статья Google ученый
Daniels, C.B. & Orgeig, S. Легочный сурфактант: ключ к эволюции дыхания воздухом. Н. Физиол. науч. 18 , 151–157 (2003).
КАС Google ученый
Фесслер, М. Б. и Саммер, Р. С. Липиды поверхностно-активных веществ на границе раздела хозяин-окружающая среда. метаболические сенсоры, супрессоры и эффекторы воспалительного заболевания легких. 900:15 утра. Дж. Дыхание. Ячейка Мол. биол. 54 , 624–635 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
 «>
«>Эррера-Рамос, Э. и др. Генетические варианты сурфактантного белка А связаны с тяжелой дыхательной недостаточностью при пандемической инфекции вируса гриппа А. Крит. Care 18 , R127 (2014).
ПабМед ПабМед Центральный Статья Google ученый
Glasser, S.W. et al. Мыши с дефицитом сурфактантного протеина С восприимчивы к инфекции респираторно-синцитиального вируса. утра. Дж. Физиол. Мол.клеток легких. Физиол. 297 , Л64–72 (2009).
КАС пабмед ПабМед Центральный Статья Google ученый
Флорос, Дж. и Томас, Нью-Джерси. Генетика белков сурфактанта при внебольничной пневмонии: уравновешивание воспалительного состояния хозяина. Крит. Уход 15 , 156 (2011).
ПабМед ПабМед Центральный Статья Google ученый
 «>
«>Vieira, F., Kung, JW & Bhatti, F. Структура, генетика и функция связанных с легкими белков сурфактанта A и D: внелегочная роль этих лектинов C-типа. Энн. Анат. 211 , 184–201 (2017).
ПабМед ПабМед Центральный Статья Google ученый
Лет-Ларсен Р., Чжун Ф., Чоу В. Т., Холмсков У. и Лу Дж. Гликопротеин спайка коронавируса SARS избирательно распознается белком сурфактанта легких D и активирует макрофаги. Иммунобиология 212 , 201–211 (2007).
КАС пабмед ПабМед Центральный Статья Google ученый
Woods, P. S. et al. Летальная инфекция вируса гриппа А h2N1 изменяет липидом сурфактанта мышиных альвеолярных клеток типа II. 900:15 утра. Дж. Физиол. Мол.клеток легких. Физиол. 311 , L1160–L1169 (2016).
ПабМед ПабМед Центральный Статья Google ученый
 «>
«>Sun, X. et al. Метаболизм жирных кислот связан с тяжестью заболевания после инфицирования H7N9. EBioMedicine 33 , 218–229 (2018).
ПабМед ПабМед Центральный Статья Google ученый
Окс, М. Чем ближе мы смотрим, тем больше видим? Количественный микроскопический анализ легочной сурфактантной системы. Клеточная физиол. Биохим. 25 , 27–40 (2010).
КАС пабмед Статья Google ученый
Grathwohl, C., Newman, G.E., Phizackerley, PJ & Town, MH. Структурные исследования пластинчатых осмиофильных телец, выделенных из легких свиней. Результаты ЯМР 31P и содержание воды. Биохим. Биофиз. Acta 552 , 509–518 (1979).
КАС пабмед Статья Google ученый
Voorhout, WF, Weaver, T.
 E., Haagsman, H.P., Geuze, HJ & Van Golde, LM. Биосинтетическая маршрутизация белков легочного сурфактанта в альвеолярных клетках типа II. Микроск. Рез. Тех. 26 , 366–373 (1993).
E., Haagsman, H.P., Geuze, HJ & Van Golde, LM. Биосинтетическая маршрутизация белков легочного сурфактанта в альвеолярных клетках типа II. Микроск. Рез. Тех. 26 , 366–373 (1993).КАС пабмед Статья Google ученый
Hobi, N. et al. Маленький ключик открывает тяжелую дверь: основная функция малых гидрофобных белков SP-B и SP-C заключается в том, чтобы инициировать адсорбцию пластинчатых тел легочного сурфактанта. Биохим. Биофиз. Acta 1863 , 2124–2134 (2016).
КАС пабмед Статья Google ученый
Мулугета, С. и др. Идентификация LBM180, мембранного белка, ограничивающего пластинчатые тела альвеолярных клеток типа II, в качестве белка-транспортера ABC ABCA3. J. Biol. хим. 277 , 22147–22155 (2002 г.).
КАС пабмед Статья Google ученый
 «>
«>Мацумура, Ю., Сакаи, Х., Сасаки, М., Бан, Н. и Инагаки, Н. Опосредованное ABCA3 поглощение холин-фосфолипидов внутриклеточными везикулами в клетках A549. ФЭБС Письмо. 581 , 3139–3144 (2007).
КАС пабмед Статья Google ученый
Перес-Гил, Дж. Структура мембран и пленок легочного сурфактанта: роль белков и липид-белковых взаимодействий. Биохим. Биофиз. Acta 1778 , 1676–1695 (2008 г.).
КАС пабмед Статья Google ученый
Vanhecke, D. et al. Новый взгляд на ультраструктуру пластинчатого тела: замораживание под высоким давлением и криоэлектронная микроскопия срезов стекловидного тела. Гистохим. Клеточная биол. 134 , 319–326 (2010).
КАС пабмед Статья Google ученый
 «>
«>Al-Amoudi, A., Studer, D. & Dubochet, J. Артефакты резки и процесс резки в срезах стекловидного тела для криоэлектронной микроскопии. Дж. Структура. биол. 150 , 109–121 (2005).
КАС пабмед Статья Google ученый
Ригорт, А. и др. Микрообработка сфокусированным ионным пучком эукариотических клеток для криоэлектронной томографии. Проц. Натл акад. науч. США 109 , 4449–4454 (2012).
КАС пабмед Статья ПабМед Центральный Google ученый
Marko, M., Hsieh, C., Schalek, R., Frank, J. & Mannella, C. Истончение замороженных гидратированных биологических образцов с помощью сфокусированного ионного пучка для криоэлектронной микроскопии. Нац. Методы 4 , 215–217 (2007).
КАС пабмед Статья Google ученый
 «>
«>Kukulski, W. et al. Коррелированная флуоресцентная и трехмерная электронная микроскопия с высокой чувствительностью и пространственной точностью. J. Cell Biol. 192 , 111–119 (2011).
КАС пабмед ПабМед Центральный Статья Google ученый
Schorb, M. & Briggs, J.A. Коррелированная криофлуоресцентная и криоэлектронная микроскопия с высокой пространственной точностью и улучшенной чувствительностью. Ультрамикроскопия 143 , 24–32 (2014).
КАС пабмед Статья Google ученый
Метскас, Л. А. и Бриггс, Дж. А. Г. Флуоресцентное обнаружение состояния слияния мембран на крио-ЭМ сетке с использованием коррелированной криофлуоресцентной и криоэлектронной микроскопии. Микроск. Микроанал. 25 , 942–949 (2019).
КАС пабмед ПабМед Центральный Статья Google ученый
 «>
«>Сартори-Рупп, А. и др. Корреляционная криоэлектронная микроскопия выявляет структуру ТНТ в нейрональных клетках. Нац. коммун. 10 , 342 (2019).
КАС пабмед ПабМед Центральный Статья Google ученый
Плицко Дж. М., Ригорт А. и Лейс А. Корреляционная криосветовая микроскопия и криоэлектронная томография: от клеточных территорий к молекулярным ландшафтам. Курс. мнение Биотехнолог. 20 , 83–89 (2009).
КАС пабмед Статья Google ученый
Schellenberger, P. et al. Высокоточная корреляционная флуоресцентная и электронная криомикроскопия с использованием двух независимых маркеров совмещения. Ультрамикроскопия 143 , 41–51 (2014).
КАС пабмед ПабМед Центральный Статья Google ученый
 «>
«>Hampton, C.M. et al. Коррелированная флуоресцентная микроскопия и криоэлектронная томография инфицированных или трансфицированных вирусом клеток млекопитающих. Нац. протокол 12 , 150–167 (2017).
КАС пабмед Статья Google ученый
Арнольд Дж. и др. Подготовка образца криофокусированным ионным пучком для конкретного места под контролем 3D коррелятивной микроскопии. Биофиз. J. 110 , 860–869 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Wilfling, F. et al. Селективный путь аутофагии для разделенных фаз эндоцитарных белковых отложений. Мол. Клетка. 80 , 764–778.e7 (2020).
КАС пабмед ПабМед Центральный Статья Google ученый
Либер, М.
 , Смит, Б., Сзакал, А., Нельсон-Риз, В. и Тодаро, Г. Непрерывная линия опухолевых клеток карциномы легких человека со свойствами клеток альвеолярного эпителия II типа. Междунар. Дж. Рак 17 , 62–70 (1976).
, Смит, Б., Сзакал, А., Нельсон-Риз, В. и Тодаро, Г. Непрерывная линия опухолевых клеток карциномы легких человека со свойствами клеток альвеолярного эпителия II типа. Междунар. Дж. Рак 17 , 62–70 (1976).КАС пабмед Статья Google ученый
Wolff, G. et al. Обратите внимание на зазор: микрокомпенсационные швы резко уменьшают изгиб криопластин, отфрезерованных FIB. Дж. Структура. биол. 208 , 107389 (2019).
ПабМед Статья Google ученый
Castano-Diez, D., Kudryashev, M., Arheit, M. & Stahlberg, H. Dynamo: гибкий и удобный инструмент разработки для усреднения субтомограмм крио-ЭМ данных в высокопроизводительных вычислительных средах. Дж. Структура. биол. 178 , 139–151 (2012).
ПабМед Статья Google ученый
 «>
«>Cooper, J. R. et al. Длительное культивирование линии раковых клеток A549 способствует формированию многослойных телец и дифференцировке в сторону фенотипа альвеолярных пневмоцитов II типа. PLoS ONE 11 , e0164438 (2016).
ПабМед ПабМед Центральный Статья КАС Google ученый
Горелик, С. и др. PIE-scope, встроенный криокорреляционный свет и микроскопия FIB/SEM. Элиф https://doi.org/10.7554/eLife.45919 (2019).
Кришнамурти Г., Ван С.Ю., Стейер Г. и Уилсон Д.Л. Удаление подповерхностной флуоресценции при криовизуализации с использованием деконволюции. Опц. Экспресс 18 , 22324–22338 (2010 г.).
ПабМед ПабМед Центральный Статья Google ученый
Faoro, R. et al. Криоиммерсионная световая микроскопия с коррекцией аберраций.
 Проц. Натл акад. науч. США 115 , 1204–1209 (2018).
Проц. Натл акад. науч. США 115 , 1204–1209 (2018).КАС пабмед Статья ПабМед Центральный Google ученый
Вольф Г., Хаген К., Грюневальд К. и Кауфманн Р. На пути к корреляционной флуоресценции сверхвысокого разрешения и электронной криомикроскопии. Биол. Cell 108 , 245–258 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Haller, T., Cerrada, A., Pfaller, K., Braubach, P. & Felder, E. Микроскопия в поляризованном свете выявляет физиологические и лекарственные изменения в сборке мембран сурфактанта в альвеолярных пневмоцитах II типа. Биохим. Биофиз. Акта Биомембр. 1860 , 1152–1161 (2018).
КАС пабмед Статья Google ученый
Махамид, Дж.
 и др. Жидкокристаллические фазовые переходы в каплях липидов связаны с клеточными состояниями и специфической ассоциацией органелл. Проц. Натл акад. науч. США 116 , 16866–16871 (2019).
и др. Жидкокристаллические фазовые переходы в каплях липидов связаны с клеточными состояниями и специфической ассоциацией органелл. Проц. Натл акад. науч. США 116 , 16866–16871 (2019).КАС пабмед Статья ПабМед Центральный Google ученый
Engelbrecht, S., Kaltenborn, E., Griese, M. & Kern, S. Переносчик липидов сурфактанта ABCA3 расщеплен на N-конце внутри LAMP3-позитивных везикул. ФЭБС Письмо. 584 , 4306–4312 (2010).
КАС пабмед Статья Google ученый
Бхоумик, Р. и др. Трехмерная модель легкого человека, созданная с помощью тканевой инженерии, для изучения инфекции гриппа А. Ткань Eng. Часть A 24 , 1468–1480 (2018).
КАС пабмед ПабМед Центральный Статья Google ученый
 «>
«>Чжан, Л. и др. Rab38 нацеливается на ламеллярные тельца и нормализует их размеры в эпителиальных клетках альвеолярного типа II легких. утра. Дж. Физиол. Мол.клеток легких. Физиол. 301 , L461–477 (2011).
КАС пабмед ПабМед Центральный Статья Google ученый
Танака Х. и др. Структура свода печени крысы при разрешении 3,5 ангстрем. Наука 323 , 384–388 (2009).
КАС пабмед Статья Google ученый
Раст А. и др. Биогенные участки тилакоидов цианобактерий образуют места контакта с плазматической мембраной. Нац. Растения 5 , 436–446 (2019).
КАС пабмед Статья Google ученый
Preibisch, S., Saalfeld, S. & Tomancak, P. Глобально оптимальное сшивание мозаичных 3D микроскопических изображений.
 Биоинформатика 25 , 1463–1465 (2009).
Биоинформатика 25 , 1463–1465 (2009).КАС пабмед ПабМед Центральный Статья Google ученый
Schindelin, J. et al. Фиджи: платформа с открытым исходным кодом для анализа биологических изображений. Нац. Методы 9 , 676–682 (2012).
КАС Статья пабмед Google ученый
Мастронард, Д. Н. Автоматизированная томография с помощью электронного микроскопа с использованием надежного прогнозирования движений образца. Дж. Структура. биол. 152 , 36–51 (2005).
Артикул пабмед Google ученый
Хаген, У. Дж. Х., Ван, У. и Бриггс, Дж. А. Г. Реализация схемы наклона криоэлектронной томографии, оптимизированной для усреднения субтомограмм с высоким разрешением. Дж. Структура.
 биол. 197 , 191–198 (2017).
биол. 197 , 191–198 (2017).ПабМед ПабМед Центральный Статья Google ученый
Мастронард, Д. Н. и Хелд, С. Р. Автоматическое выравнивание серии наклонов и томографическая реконструкция в IMOD. Дж. Структура. биол. 197 , 102–113 (2017).
ПабМед Статья Google ученый
Paul-Gilloteaux, P. et al. eC-CLEM: гибкое программное обеспечение для многомерной регистрации для коррелятивной микроскопии. Нац. Методы 14 , 102–103 (2017).
КАС пабмед Статья Google ученый
Загрузить ссылки
Благодарности
Мы хотели бы поблагодарить д-ра Surafel Mulugeta за любезно предоставленную плазмиду hABCA3-eGFP и проф. д-р Hans-Georg Kräuslich за предоставление клеток HSAEpC. Мы благодарим доктора Стефана Пфеффера за критическое прочтение рукописи и докторов Бена Энгеля и доктора Ларса Андерса Карлсона за плодотворное обсуждение OMDP. Мы хотели бы поблагодарить Платформу визуализации инфекционных заболеваний (IDIP) в Центре интегративных исследований инфекционных заболеваний Гейдельберга за поддержку в области микроскопии. Мы хотели бы подтвердить доступ к инфраструктуре и поддержке, предоставляемой сетью Cryo-EM в Гейдельбергском университете (HDcryoNet). Это исследование финансировалось за счет исследовательского гранта Фонда Чики и Хайнца Шаллеров (Программа руководителя исследовательской группы Шаллера) и Немецкого исследовательского общества (DFG, Немецкий исследовательский фонд) Номер проекта 240245660 SFB 1129..
Мы хотели бы поблагодарить Платформу визуализации инфекционных заболеваний (IDIP) в Центре интегративных исследований инфекционных заболеваний Гейдельберга за поддержку в области микроскопии. Мы хотели бы подтвердить доступ к инфраструктуре и поддержке, предоставляемой сетью Cryo-EM в Гейдельбергском университете (HDcryoNet). Это исследование финансировалось за счет исследовательского гранта Фонда Чики и Хайнца Шаллеров (Программа руководителя исследовательской группы Шаллера) и Немецкого исследовательского общества (DFG, Немецкий исследовательский фонд) Номер проекта 240245660 SFB 1129..
Финансирование
Финансирование открытого доступа организовано и разрешено Projekt DEAL.
Информация об авторе
Примечания автора
Эти авторы внесли равный вклад: Штеффен Кляйн, Бенедикт Х. Виммер.
Авторы и филиалы
Schaller Research Groups, Кафедра инфекционных болезней и вирусологии, Гейдельбергский университет, Гейдельберг, Германия
Штеффен Кляйн, Бенедикт Х.
 Виммер, Софи Колан99, Колланд Винтеру, Андрон0005
Виммер, Софи Колан99, Колланд Винтеру, Андрон0005Исследовательская группа «Мембранная биология вирусных инфекций», кафедра инфекционных болезней и вирусологии Гейдельбергского университета, Гейдельберг, Германия
Штеффен Кляйн, Бенедикт Х. Виммер, Софи Л. Винтер, Андроники Колову и Петр Хланда
Infectious Diseases Imaging Platform (IDIP), Кафедра инфекционных болезней Гейдельбергского университета, Гейдельберг, Германия
Vibor Laketa
Авторы
- Steffen Klein
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Benedikt H. Wimmer
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Sophie L. Winter
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Андроники Коловой
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Vibor Laketa
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Петр Хланда
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Взносы
Подготовка проб: S. K., B.H.W., S.L.W., A.K. и PC; исследование концептуализации и дизайна: С.К., Б.Х.В., В.Л. и П.К.; сбор данных: S.K., B.H.W., S.L.W., A.K. и PC; анализ: С.К., Б.Х.В., С.Л.В., В.Л. и П.К., подготовка бумаги: С.К., Б.Х.В., С.Л.В., и П.К.
K., B.H.W., S.L.W., A.K. и PC; исследование концептуализации и дизайна: С.К., Б.Х.В., В.Л. и П.К.; сбор данных: S.K., B.H.W., S.L.W., A.K. и PC; анализ: С.К., Б.Х.В., С.Л.В., В.Л. и П.К., подготовка бумаги: С.К., Б.Х.В., С.Л.В., и П.К.
Автор, ответственный за переписку
Петр Хланда.
Заявление об этике
Конкурирующие интересы
Авторы не заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Примечание издателя Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Дополнительная информация
Дополнительная информация
Описания дополнительных дополнительных файлов
Дополнительный фильм 1
Дополнительный фильм 2
Дополнительный фильм 3 9.0201
Дополнительные данные 1
Резюме отчета
Права и разрешения
Открытый доступ любой носитель или формат, при условии, что вы укажете автора(ов) оригинала и источник, предоставите ссылку на лицензию Creative Commons и укажете, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или выходит за рамки разрешенного использования, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или выходит за рамки разрешенного использования, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатка и разрешения
Об этой статье
Дополнительная литература
Микровезикулы, высвобождаемые из эпителиальных клеток легких, стимулированных пневмолизином, несут митохондриальный груз и подавляют окислительный взрыв нейтрофилов.
- Э.
 Летсиу
Летсиу - Л. Г. Тейшейра Алвес
- М. Витценрат
Научные отчеты (2021)
- Э.
Комментарии
Отправляя комментарий, вы соглашаетесь соблюдать наши Условия и Правила сообщества. Если вы обнаружите что-то оскорбительное или не соответствующее нашим условиям или правилам, отметьте это как неприемлемое.
Что такое ламеллярная вода в продуктах для волос?
«Жидкие волосы» сейчас повсюду: на красных дорожках, в Инстаграме и в аптеке. Возможно, именно поэтому такие продукты, как Garnier Sleek & Shine Glass Hair Water (8 долларов) и L’Oréal Paris Elvive Dream Lengths Wonder Water (9 долларов). ) становятся все более популярными в прошлом году, и люди, испробовавшие лечение, часто называют ее «чудодейственной водой». Причина, по которой они настолько эффективны, кроется в так называемой ламеллярной воде. Но в то время как все совершенно одержимы продуктами (и полученным внешним видом), никто на самом деле не знает, что такое ламеллярная вода на самом деле — кроме косметических химиков.
) становятся все более популярными в прошлом году, и люди, испробовавшие лечение, часто называют ее «чудодейственной водой». Причина, по которой они настолько эффективны, кроется в так называемой ламеллярной воде. Но в то время как все совершенно одержимы продуктами (и полученным внешним видом), никто на самом деле не знает, что такое ламеллярная вода на самом деле — кроме косметических химиков.
Провозглашенный новейшим ингредиентом для волос, мы хотели точно знать, что делает его таким особенным. Мы попросили химика-косметолога Эстер Олу ответить на наши острые вопросы о технологии и о том, соответствует ли ламеллярная вода шумихе, наводнившей наши ленты в Instagram и TikTok. Продолжайте читать, чтобы узнать.
Что такое ламеллярная вода?
Ламеллярная вода — это смываемое кондиционирующее и восстанавливающее средство для волос, запатентованное L’Oréal Paris, что означает, что это единственный косметический конгломерат с ламеллярной технологией. И эта технология была впервые представлена в 2019 году компанией Kérastase (принадлежит L’Oréal) в виде салонного средства под названием K Water, которое, как и новая чудо-вода L’Oréal Paris, обещало «восстанавливать волокна волос и придает волосам гладкую поверхность», что делает его фаворитом в салонах.
Перенесемся в сегодняшний день: технология была разработана для безопасного и эффективного лечения в домашних условиях, что делает ее более доступной и доступной. Несколько ламеллярных средств для ухода за водой, доступных прямо сейчас, включают вышеупомянутые L’Oréal Paris Wonder Water и Garnier Sleek & Shine, а также ламеллярное средство для очистки воды Extreme Bleach Recovery от Redken (27 долларов США), все из которых имеют очень похожую технологию, но предназначены для немного другие проблемы с волосами. «Я действительно думаю, что разница в том, как они продаются», — говорит Олу. «В формулу Redken могли быть добавлены дополнительные активные вещества для лечения обесцвеченных волос, но я уверен, что оба они будут лечить волосы очень похожим образом».
Как работает ламеллярная вода?
«Ламеллярная вода — это технология, которая нацелена на поврежденные участки волос путем нанесения концентрированных активных ингредиентов, таких как аминокислоты и белки, которые полезны для восстановления прядей», — говорит Олу. Как и следовало ожидать, ламеллярная вода очень легкая, как и вода из-под крана.
Как и следовало ожидать, ламеллярная вода очень легкая, как и вода из-под крана.
Именно это делает лечение таким особенным. «Активы волос распределяются очень тонкими слоями, называемыми ламелями, что позволяет им оседать на волосах и проникать в кутикулу волоса», — говорит Олу. «Вместо того, чтобы покрывать всю прядь волос, ламеллярная вода предназначена для воздействия на те области, где она необходима и где происходит повреждение».
«Все дело в положительных и отрицательных зарядах, — говорит старший научный консультант L’Oréal UK&I Керис Маллен. «Ламели — это то, как устроена жидкость Wonder Water; она состоит из различных слоев. Это работает, потому что она может упаковывать кондиционеры в меньшее пространство, поэтому тонкий слой кондиционеров доставляется к стержню волоса. Поврежденные участки — это другой заряд. к кондиционеру, и поэтому [кондиционер] притягивается к поврежденным участкам и интенсивно воздействует на них».
Чем ламеллярная вода отличается от кондиционера?
Вам может быть интересно, есть ли большая разница между пластинчатой обработкой воды и кондиционерами. Короткий ответ: да, они, безусловно, разные, когда дело доходит до составов. «Основное различие между ламеллярной водой и кондиционерами заключается в том, что ламеллярная вода нацелена на поврежденные части волос, а не на всю прядь волос, где волосы считаются здоровыми и неповрежденными; обычно это корни», — говорит Олу. Покрывая ламеллярную воду только поврежденную часть прядей волос и доставляя активные ингредиенты непосредственно в эти области, он не утяжеляет волосы, воздействуя только на области, нуждающиеся в дополнительной ТСХ.
Короткий ответ: да, они, безусловно, разные, когда дело доходит до составов. «Основное различие между ламеллярной водой и кондиционерами заключается в том, что ламеллярная вода нацелена на поврежденные части волос, а не на всю прядь волос, где волосы считаются здоровыми и неповрежденными; обычно это корни», — говорит Олу. Покрывая ламеллярную воду только поврежденную часть прядей волос и доставляя активные ингредиенты непосредственно в эти области, он не утяжеляет волосы, воздействуя только на области, нуждающиеся в дополнительной ТСХ.
, с другой стороны, имеют гораздо более густую консистенцию, чем ламеллярная вода. По словам Олу, из-за этой более высокой вязкости, в дополнение к добавлению таких ингредиентов, как силиконы, традиционные кондиционеры могут иногда покрывать волосы, делая их тяжелыми или отягощенными. Как ламеллярная вода, так и традиционные средства для кондиционирования технически все еще считаются кондиционерами, но ламеллярная вода намного легче, что делает ее особенно отличным вариантом, если вы обнаружите, что традиционные кондиционеры делают ваши волосы тяжелыми, утяжеленными или жирными.
Подходит ли ламеллярная вода для всех типов волос?
L’Oréal Paris утверждает, что ламеллярная чудо-вода подходит для всех типов волос: тонких, густых, вьющихся и вьющихся. «Мы протестировали чудо-воду Elvive Dream Lengths на всех типах волос, включая афро-волосы, и наши результаты подтвердили, что она работает на всех типах волос», — говорит Маллен. «Он идеально подходит для людей с сухими, ломкими или поврежденными волосами, которые могут возникнуть в результате окрашивания, обесцвечивания, чрезмерной укладки или теплового повреждения», — говорит Олу.
Как использовать ламеллярную воду
Как и любой традиционный кондиционер или маску, ламеллярную воду следует наносить на середину и кончики волос после мытья шампунем. Подождав несколько секунд, вы просто смоете, и все готово. Поскольку ламеллярная вода, очевидно, имеет водную основу, она не имеет кремообразной консистенции, к которой вы, вероятно, привыкли от обычного кондиционера; однако пенится очень слабо.
Вот оно. Довольно простая формула ламеллярной воды на самом деле невероятно трудоемка и может стать вашим билетом к кондиционированным и восстановленным волосам, не утяжеляя их. Более того, что касается ухода за волосами, ламеллярные водные процедуры довольно доступны. Еще один момент, на который стоит обратить внимание, это запах. Запах сильный, действительно приятный фруктовый запах, но если вы чувствительны к ароматам в своих продуктах, об этом стоит помнить.
Источник изображения: Getty/RB/Bauer-Griffin
Секреторная реакция пластинчатых телец на нарушение барьера
. 1992 март; 98 (3): 279-89.
дои: 10.1111/1523-1747.ep12497866.
Г К Менон 1 , К. Р. Фейнгольд, П. М. Элиас
Р. Фейнгольд, П. М. Элиас
принадлежность
- 1 Дерматологическая служба, Медицинский центр Управления по делам ветеранов, Сан-Франциско, Калифорния 94121.
- PMID: 1545137
- DOI: 10.1111/1523-1747.ep12497866
Бесплатная статья
GK Menon et al. Джей Инвест Дерматол. 1992 март
Бесплатная статья
. 1992 март; 98 (3): 279-89.
дои: 10. 1111/1523-1747.ep12497866.
1111/1523-1747.ep12497866.
Авторы
Г К Менон 1 , К. Р. Фейнгольд, П. М. Элиас
принадлежность
- 1 Дерматологическая служба, Медицинский центр Управления по делам ветеранов, Сан-Франциско, Калифорния 94121.
- PMID: 1545137
- DOI: 10.1111/1523-1747.ep12497866
Абстрактный
Многочисленные данные указывают на важную роль секреции эпидермальных пластинчатых телец в поддержании барьера проницаемости. Однако реакция секреторной системы пластинчатых тел на разрушение барьера не исследована. Следовательно, мы исследовали секреторную реакцию пластинчатых тел в различных точках после индуцированного ацетоном снятия барьера у бесшерстных мышей у животных, подвергавшихся воздействию воздуха, и у животных, окклюзированных непроницаемыми мембранами по сравнению с паропроницаемыми. Животные, лишенные изоленты, служили контролем химической токсичности. Нарушение барьера либо ацетоном, либо липкой лентой сопровождалось быстрой секрецией содержимого ламеллярных телец из самого верхнего слоя зернистых клеток, в результате чего цитозоль в значительной степени был лишен ламеллярных тел. Вновь секретируемое содержимое ламеллярных тел состояло из складчатых пластинок (а не «дисков», как считалось ранее), которые развертывались в межклеточных пространствах на границе зернисто-ороговевших клеток. В это время (15-30 мин) основная единица структуры ламеллярных бислоев в среднем и верхнем роговом слое казалась дезорганизованной и перемежалась большими лакунами, отражающими экстракцию растворителем.
Следовательно, мы исследовали секреторную реакцию пластинчатых тел в различных точках после индуцированного ацетоном снятия барьера у бесшерстных мышей у животных, подвергавшихся воздействию воздуха, и у животных, окклюзированных непроницаемыми мембранами по сравнению с паропроницаемыми. Животные, лишенные изоленты, служили контролем химической токсичности. Нарушение барьера либо ацетоном, либо липкой лентой сопровождалось быстрой секрецией содержимого ламеллярных телец из самого верхнего слоя зернистых клеток, в результате чего цитозоль в значительной степени был лишен ламеллярных тел. Вновь секретируемое содержимое ламеллярных тел состояло из складчатых пластинок (а не «дисков», как считалось ранее), которые развертывались в межклеточных пространствах на границе зернисто-ороговевших клеток. В это время (15-30 мин) основная единица структуры ламеллярных бислоев в среднем и верхнем роговом слое казалась дезорганизованной и перемежалась большими лакунами, отражающими экстракцию растворителем. Зарождающиеся ламеллярные тельца начали вновь появляться в цитозоле зернистых клеток к 30 мин, а к 360 мин клетки демонстрировали полный набор нормально выглядящих ламеллярных тел. Между 60 и 360 мин плотность ламеллярных слоев тела на границе гранулярно-ороговевших клеток увеличивалась, в то время как мембранные бислои наружного рогового слоя оставались дезорганизованными. Новые ламеллярные бислои сначала появлялись в нижнем роговом слое между 60 и 180 мин в результате трансформации секретируемых ламеллярных слоев тела, и со временем эти ламеллы появлялись в более апикальных местах. Окклюзия паронепроницаемой, но не паропроницаемой мембраной привела к а) уменьшению количества пластинчатых телец и межклеточных продуктов, происходящих из пластинчатых телец; б) образование пластинчатых тел с аномальным внутренним содержимым; в) угнетение секреции пластинчатых телец; и d) ингибирование превращения пластинок, происходящих из ламеллярного тела, в ламеллярные двухслойные единицы. Эти результаты демонстрируют центральную роль пластинчато-секреторной системы в восстановлении барьера и гомеостазе.
Зарождающиеся ламеллярные тельца начали вновь появляться в цитозоле зернистых клеток к 30 мин, а к 360 мин клетки демонстрировали полный набор нормально выглядящих ламеллярных тел. Между 60 и 360 мин плотность ламеллярных слоев тела на границе гранулярно-ороговевших клеток увеличивалась, в то время как мембранные бислои наружного рогового слоя оставались дезорганизованными. Новые ламеллярные бислои сначала появлялись в нижнем роговом слое между 60 и 180 мин в результате трансформации секретируемых ламеллярных слоев тела, и со временем эти ламеллы появлялись в более апикальных местах. Окклюзия паронепроницаемой, но не паропроницаемой мембраной привела к а) уменьшению количества пластинчатых телец и межклеточных продуктов, происходящих из пластинчатых телец; б) образование пластинчатых тел с аномальным внутренним содержимым; в) угнетение секреции пластинчатых телец; и d) ингибирование превращения пластинок, происходящих из ламеллярного тела, в ламеллярные двухслойные единицы. Эти результаты демонстрируют центральную роль пластинчато-секреторной системы в восстановлении барьера и гомеостазе.
Похожие статьи
Морфологические изменения ламеллярных тел и липидов интеркорнеоцитов после снятия ленты и окклюзии паронепроницаемой мембраной.
Цзян С., Ку С.В., Ли С.Х. Цзян С. и др. Арка Дерматол Рез. 1998 март; 290(3):145-51. doi: 10.1007/s004030050280. Арка Дерматол Рез. 1998. PMID: 9558490
Структурная основа аномалии барьера после ингибирования редуктазы ГМГ-КоА в эпидермисе мышей.
Менон Г.К., Фейнгольд К.Р., Мао-Цян М., Шауд М., Элиас П.М. Менон Г.К. и соавт. Джей Инвест Дерматол. 1992 г., февраль; 98 (2): 209–19. дои: 10.1111/1523-1747.ep12555880. Джей Инвест Дерматол. 1992. PMID: 1732385
Локализация кальция в эпидермисе мышей после разрушения и восстановления барьера проницаемости.
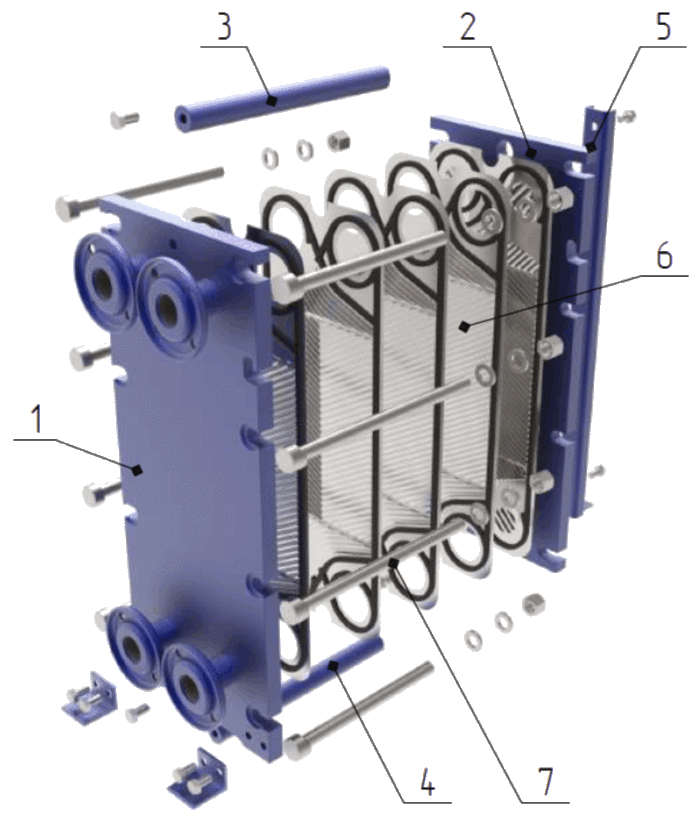
Менон Г.К., Элиас П.М., Ли С.Х., Фейнгольд К.Р. Менон Г.К. и соавт. Сотовые Ткани Res. 1992 декабрь; 270 (3): 503-12. дои: 10.1007/BF00645052. Сотовые Ткани Res. 1992. PMID: 1486603
Регуляция и роль эпидермального синтеза липидов.
Фейнгольд КР. Фейнгольд КР. Adv Липид Res. 1991;24:57-82. doi: 10.1016/b978-0-12-024924-4.50007-9. Adv Липид Res. 1991. PMID: 1763718 Обзор.
Барьерная функция регулирует синтез эпидермальных липидов и ДНК.
Прокш Э., Холлеран В.М., Менон Г.К., Элиас П.М., Фейнгольд К.Р. Прокш Э. и соавт. Бр Дж Дерматол. 1993 г., май; 128(5):473-82. doi: 10.1111/j.1365-2133.1993.tb00222.x. Бр Дж Дерматол. 1993. PMID: 8504036 Обзор.

Посмотреть все похожие статьи
Цитируется
Воспаление 2 типа способствует дисфункции кожного барьера при атопическом дерматите.
Бек Л.А., Корк М.Дж., Амагай М., Де Бенедетто А., Кабашима К., Гамильтон Д.Д., Росси А.Б. Бек Л.А. и соавт. JID Иннов. 2022 26 апреля; 2(5):100131. doi: 10.1016/j.xjidi.2022.100131. электронная коллекция 2022 сент. JID Иннов. 2022. PMID: 36059592 Бесплатная статья ЧВК. Обзор.
Кожный барьер у младенцев: обновление и перспективы.
Рахма А., Лейн М.Е. Рахма А. и др. Фармацевтика. 2022 17 февраля; 14 (2): 433. doi: 10.3390/фармацевтика14020433. Фармацевтика. 2022. PMID: 35214165 Бесплатная статья ЧВК. Обзор.

Лорикрин и NRF2 координируют ороговение.
Исицука Ю., Огава Т., Накамура Ю., Кубота Н., Фудзисава Ю., Ватанабэ Р., Окияма Н., Фудзимото М., Руп Д.Р., Исида-Ямамото А. Ишицука Ю. и др. JID Иннов. 2021 15 октября; 2(1):100065. doi: 10.1016/j.xjidi.2021.100065. Электронная коллекция 2022 янв. JID Иннов. 2021. PMID: 35024686 Бесплатная статья ЧВК.
Электрические аспекты кожи как путь к инженерным кожным устройствам.
Абэ Ю., Нисидзава М. Абэ Ю. и др. АПЛ Биоинж. 2021 18 ноября; 5 (4): 041509. дои: 10.1063/5.0064529. электронная коллекция 2021 дек. АПЛ Биоинж. 2021. PMID: 34849444 Бесплатная статья ЧВК. Обзор.
Разработка пластырей с микроиглами для улучшения проникновения: анализ, модели кожи и факторы, влияющие на введение иглы.
